

Review
. 2016 Jun;4(3):10.1128/microbiolspec.MCHD-0008-2015.
doi: 10.1128/microbiolspec.MCHD-0008-2015.
Antigen Processing and Presentation Mechanisms in Myeloid Cells
Affiliations
- PMID: 27337472
- PMCID: PMC6300129
- DOI: 10.1128/microbiolspec.MCHD-0008-2015
Item in Clipboard
Review
Antigen Processing and Presentation Mechanisms in Myeloid Cells
Microbiol Spectr.
2016 Jun.
Abstract
Unlike B cells, CD8-positive and CD4-positive T cells of the adaptive immune system do not recognize intact foreign proteins but instead recognize polypeptide fragments of potential antigens. These antigenic peptides are expressed on the surface of antigen presenting cells bound to MHC class I and MHC class II proteins. Here, we review the basics of antigen acquisition by antigen presenting cells, antigen proteolysis into polypeptide fragments, antigenic peptide binding to MHC proteins, and surface display of both MHC class I-peptide and MHC class II-peptide complexes.
Figures
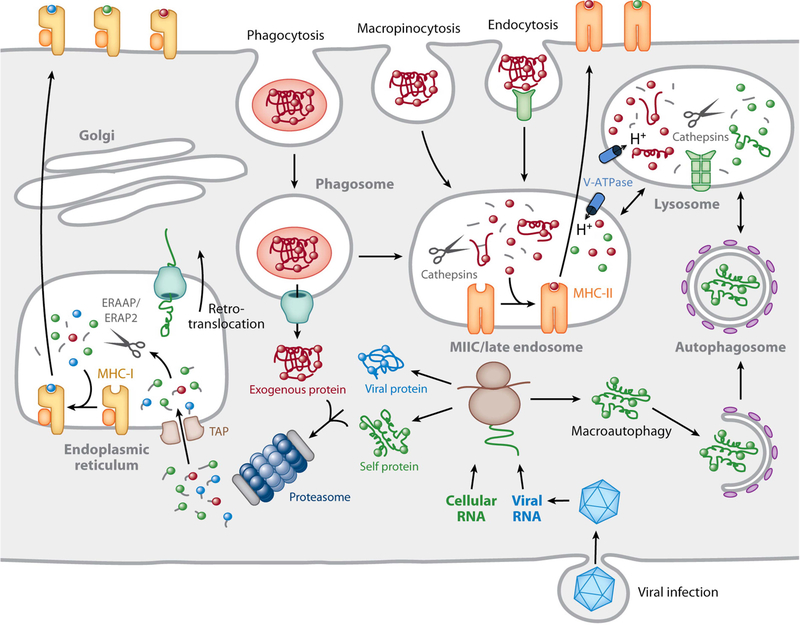
Overview of MHC-peptide complex biogenesis. Cytosolic proteins are degraded by the proteasome into small peptides that are imported into the lumen of the ER by TAP, where they bind to nascent MHC-I molecules. ER peptides can be trimmed to 8 to 10 residues by the action of ERAAP/ERAP1 and ERAP2. Fully assembled MHC-I–peptide complexes leave the ER and are delivered to the plasma membrane by recognition by CD8+ T cells. Proteins internalized into endosomes by a variety of mechanisms are degraded into peptides in late endosomes rich in proteinases, classically called cathepsins, active at acidic pH. MHC-II molecules are transported to these compartments from the ER by virtue of its association with a chaperone termed the invariant chain (not shown). The MHC-II-positive compartment is indicated as MIIC/late endosome in the figure. Invariant chain is also proteolytically degraded in late endosomes, thereby making the MHC-II molecules available for peptide binding. Following a series of peptide-editing processes, immunodominant MHC-II–peptide complexes move to the plasma membrane for recognition by CD4+ T cells. In specialized APCs, particularly DCs, proteins that enter the cell by endocytosis/phagocytosis are retrotranslocated into the cytosol for subsequent proteasomal degradation and binding to MHC-I in a process termed cross-presentation. The retrotranslocation mechanism is currently undefined, but here it is depicted as a channel responsible for ERAD that may be recruited to the phagosome from the ER. This hypothesis remains unproven. Reprinted from reference , with permission.
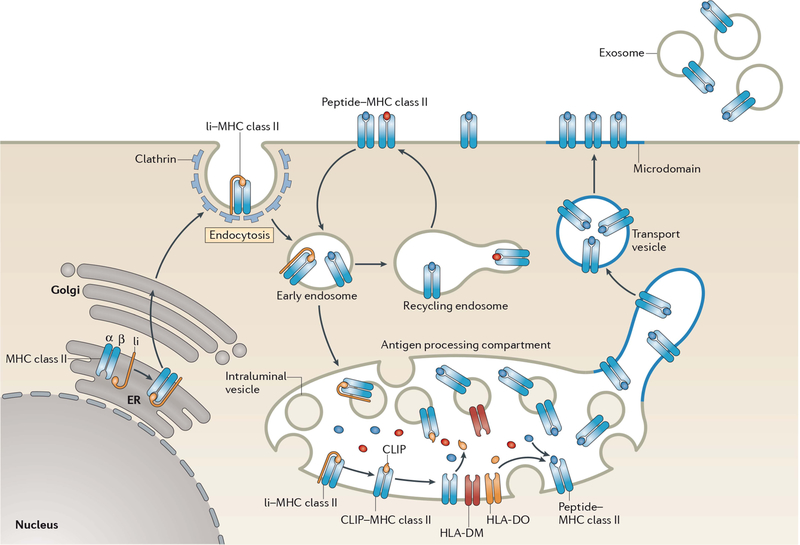
Biosynthesis of MHC-II–peptide complexes. MHC-II αβ dimers associate with Ii in the ER, and the assembled MHC-II–Ii complexes traffic through the Golgi apparatus and are delivered to the plasma membrane. The complexes are internalized by clathrin-mediated endocytosis and are transported to late endosomal multivesicular antigen-processing compartments. Some of these complexes sort onto the ILVs of these compartments, where sequential Ii proteolysis leads to persistence of a derived fragment (termed CLIP) in the MHC-II peptide-binding groove. CLIP is removed from CLIP–MHC-II complexes by DM molecules that are present on the ILV and limiting membrane of antigen-processing compartments, thereby allowing peptide binding onto nascent MHC-II. The activity of DM is regulated by DO; however, the mechanism of regulation remains unknown. It is likely that ILV-associated MHC-II is transferred to the limiting membrane and endo/lysosomal tubules that either directly fuse, or give rise to transport vesicles that fuse, with the plasma membrane. MHC-II–peptide association with lipid microdomains first occurs in antigen-processing compartments and allows clustering of MHC-II–peptide complexes on the cell surface. If an entire antigen-processing compartment fuses with the plasma membrane, the ILV can be released from the cell in the form of exosomes. Surface-expressed MHC-II–peptide complexes can internalize using a clathrin-independent endocytosis pathway and are targeted for lysosomal degradation or may be recycled back to the plasma membrane. Reprinted from reference , with permission.
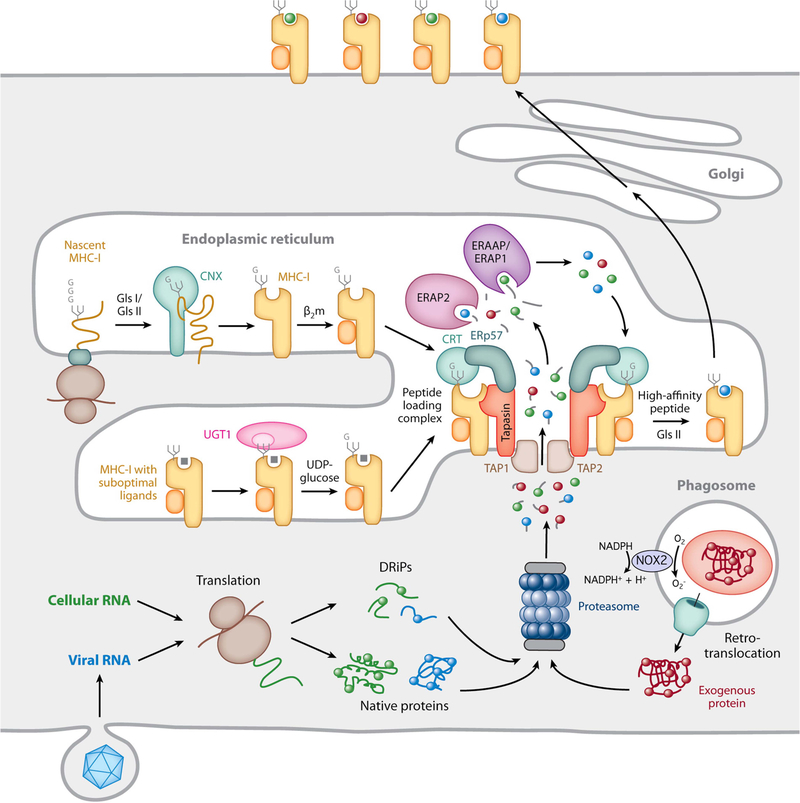
MHC-I biosynthesis and peptide binding. The proteasome generates short antigenic peptides capable of binding to MHC-I molecules. These peptides are derived from native cytosolic proteins, defective ribosomal products (DRiPs), or, in the case of cross-presentation, exogenous proteins that enter the cell by phagocytosis and are translocated into the cytosol, either intact or as large proteolytic fragments. In cross-presenting mouse CD8+ DCs, the presence of NOX2 on the phagosomal membrane neutralizes acidification and reduces proteolytic activity, preserving protein integrity. Nascent MHC-I heavy chains initially interact with the molecular chaperone calnexin (CNX) and, after binding β2m, are recruited to the PLC by simultaneous noncovalent CRT interactions with a monoglucosylated N-linked glycan on the heavy chain and ERp57 disulfide linked to tapasin in the PLC. Peptide-free MHC-I molecules and those possessing suboptimal ligands are subject to a series of “editing” steps mediated by interaction with tapasin within the PLC as well as maintenance of the monoglucosylated N-linked glycan by the opposing actions of the enzymes glucosidase 2 (GlsII), which removes the terminal glucose residue, and UGT1, which adds back glucose to preserve the CRT interaction. MHC-I molecules containing high-affinity peptides ultimately leave the ER and are transported to the plasma membrane. Reprinted from reference , with permission.
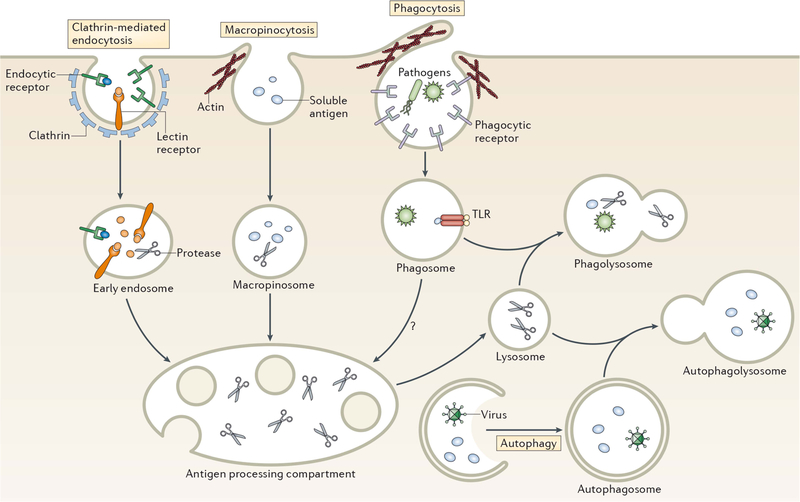
Pathways of antigen entry into the processing compartments of myeloid cells. Pathogens as well as soluble and particulate antigens access the endolysosomal pathway of antigen-processing cells by a variety of mechanisms. Clathrin-mediated endocytosis generally involves the binding of ligands to one of a variety of endocytic receptors that deliver endocytosed cargo to early endosomes. Macropinocytosis is a nonspecific form of endocytosis that involves actin-dependent membrane ruffling that leads to solute encapsulation in structures that give rise to macropinosomes. Like early endosomes, macropinosomes are not highly proteolytic and antigen degradation only occurs following their fusion with acidic late endosomal/lysosomal compartments containing lysosomal proteinases. Pathogens and large particles that possess specific binding sites for surface receptors are internalized by phagocytosis, an endocytic process that combines the features of macropinocytosis and receptor-mediated endocytosis. Phagosomes are not acidic nor proteinase rich; however, maturation of phagosomes by fusion with late endosomes or lysosomes gives rise to proteolytic phagolysosomes that degrade phagocytosed material. Autophagy also provides material for endolysosomal degradation by sequestering cytosol into a double-membrane encapsulated autophagosome that, like a conventional phagosome, undergoes maturation upon fusion with lysosomes to generate proteolytic autophagolysosomes. Reprinted from reference , with permission.
References
-
- Roche PA, Cresswell P. 1990. Invariant chain association with HLA-DR molecules inhibits immunogenic peptide binding. Nature 345:615–618. - PubMed
-
- Teyton L, O’Sullivan D, Dickson PW, Lotteau V, Sette A, Fink P, Peterson PA. 1990. Invariant chain distinguishes between the exogenous and endogenous antigen presentation pathways. Nature 348:39–44. - PubMed
-
- Bakke O, Dobberstein B. 1990. MHC class II-associated invariant chain contains a sorting signal for endosomal compartments. Cell 63:707–716. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
