

Presynaptic muscarinic acetylcholine autoreceptors (M1, M2 and M4 subtypes), adenosine receptors (A1 and A2A) and tropomyosin-related kinase B receptor (TrkB) modulate the developmental synapse elimination process at the neuromuscular junction
- PMID: 27339059
- PMCID: PMC4917939
- DOI: 10.1186/s13041-016-0248-9
Presynaptic muscarinic acetylcholine autoreceptors (M1, M2 and M4 subtypes), adenosine receptors (A1 and A2A) and tropomyosin-related kinase B receptor (TrkB) modulate the developmental synapse elimination process at the neuromuscular junction
Abstract
Background: The development of the nervous system involves an initially exuberant production of neurons that make an excessive number of synaptic contacts. The initial overproduction of synapses promotes connectivity. Hebbian competition between axons with different activities (the least active are punished) leads to the loss of roughly half of the overproduced elements and this refines connectivity and increases specificity. The neuromuscular junction is innervated by a single axon at the end of the synapse elimination process and, because of its relative simplicity, has long been used as a model for studying the general principles of synapse development. The involvement of the presynaptic muscarinic ACh autoreceptors may allow for the direct competitive interaction between nerve endings through differential activity-dependent acetylcholine release in the synaptic cleft. Then, the most active ending may directly punish the less active ones. Our previous results indicate the existence in the weakest axons on the polyinnervated neonatal NMJ of an ACh release inhibition mechanism based on mAChR coupled to protein kinase C and voltage-dependent calcium channels. We suggest that this mechanism plays a role in the elimination of redundant neonatal synapses.
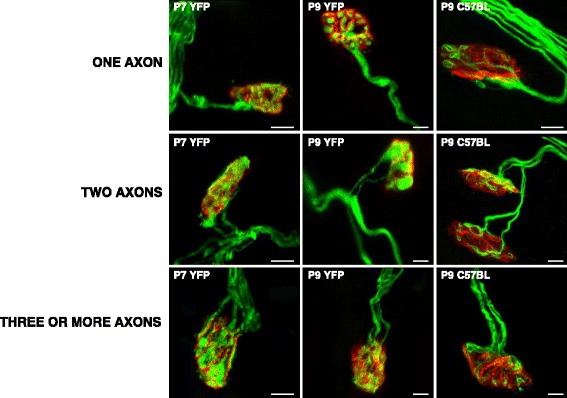
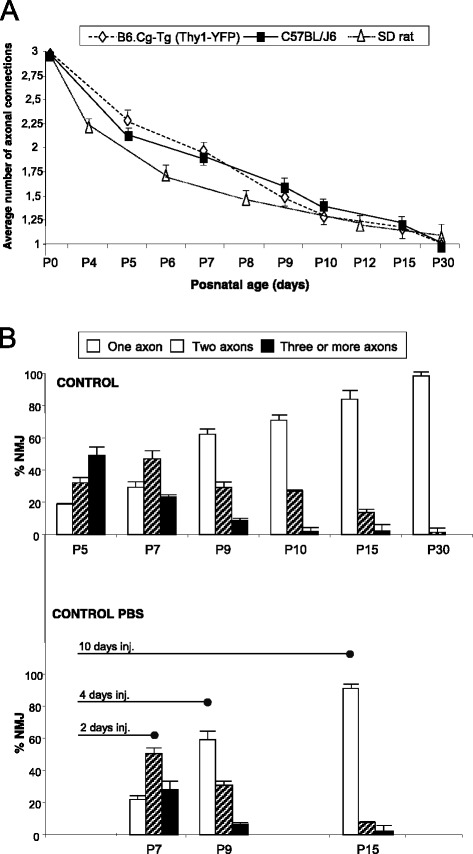
Results: Here we used confocal microscopy and quantitative morphological analysis to count the number of brightly fluorescent axons per endplate in P7, P9 and P15 transgenic B6.Cg-Tg (Thy1-YFP)16 Jrs/J mice. We investigate the involvement of individual mAChR M1-, M2- and M4-subtypes in the control of axonal elimination after the Levator auris longus muscle had been exposed to agonist and antagonist in vivo. We also analysed the role of adenosine receptor subtypes (A1 and A2A) and the tropomyosin-related kinase B receptor. The data show that postnatal axonal elimination is a regulated multireceptor mechanism that guaranteed the monoinnervation of the neuromuscular synapses.
Conclusion: The three receptor sets considered (mAChR, AR and TrkB receptors) intervene in modulating the conditions of the competition between nerve endings, possibly helping to determine the winner or the lossers but, thereafter, the final elimination would occur with some autonomy and independently of postsynaptic maturation.
Keywords: Cholinergic synapses; Motor end-plate; Motor nerve terminal; Neuromuscular junction.
Figures








References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
