

Review
doi: 10.1097/j.pain.0000000000000620.
Spotlight on pain: optogenetic approaches for interrogating somatosensory circuits
Affiliations
- PMID: 27340912
- PMCID: PMC5069102
- DOI: 10.1097/j.pain.0000000000000620
Item in Clipboard
Review
Spotlight on pain: optogenetic approaches for interrogating somatosensory circuits
Pain.
2016 Nov.
No abstract available
Conflict of interest statement
RWG is a co-founder and stockholder of Neurolux Systems. The other authors do not have a conflict of interest.
Figures
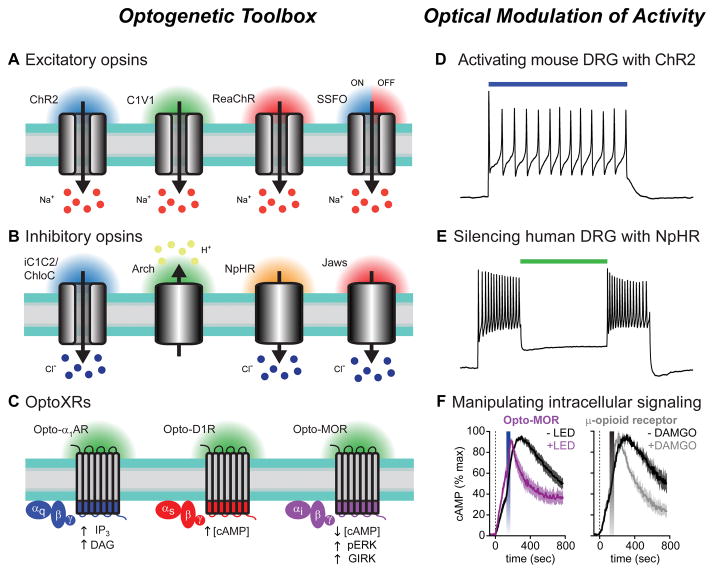
(A–C) Opsins are illustrated with their respective wavelengths of light activation. Arrows indicate direction of ion flux or effect on intracellular signaling pathways. (A) Excitatory opsins like ChR2 (far left) are nonspecific cation channels that depolarize neurons when stimulated with blue light. Interest in separately controlling different cell-types led to the development of opsins with shifted spectral properties, including the green light activatable C1V1 chimera (center left), and a red-light activatable ChR2 variant, ReaChR (center right). Stable step-function opsins (SSFOs) gate long-lasting photocurrents after brief pulses of blue light that can be rapidly terminated by red light (far right). Sodium ions flowing down their electrochemical gradient are depicted as the predominant charge carrier; however, these channels are also permeable to protons, potassium, and calcium ions. (B) iC1C2 and ChloC (far left) are channelrhodopsin variants engineered to function as inhibitory chloride-conducting channels. These opsins silence neuronal activity when illuminated with blue light. Archaerhodopsin (Arch, center left) is a green-light activated proton pump that generates proton efflux to hyperpolarize neurons. Halorhodopsins, like NpHR (center right) and Jaws (far right), are chloride pumps activated by yellow and red light, respectively. Owing to the greater tissue penetration of red-shifted light, Jaws can silence activity in vivo in combination with non-invasive transcranial illumination with red light. (C) Cartoon depicting optoXR strategy for optogenetic control of G-protein coupled receptor (GPCR) signaling. These chimeric proteins consist of the extracellular and transmembrane domains of light-sensitive rhodopsins with the intracellular regions of a GPCR of interest. Shown are Gq-coupled α1 adrenergic receptors (left, blue), Gs-coupled D1 dopamine receptors (middle, red) and Gi-coupled μ-opioid receptors (right, purple). The intracellular signaling molecules engaged by optical stimulation with blue-green light are shown below each receptor. (D) Voltage trace showing action potential firing in ChR2-expressing mouse DRG neurons during illumination with blue light (indicated by the colored bar). Modified from reference [122] with permission. (E) Using green light to silence human DRG neurons expressing the inhibitory chloride pump NpHR. Primary cultures of human sensory neurons were infected with Herpes simplex virus carrying the inhibitory opsin NpHR. Voltage trace showing sustained action potential firing during depolarizing current injections, which was inhibited when neurons were illuminated with green light (colored bar). (F) Summary graphs showing Gi-mediated inhibition of forskolin-induced cAMP production in cells expressing the light sensitive μ-opioid receptor chimera opto-MOR after blue LED stimulation (left graph, blue bar). The kinetics of cAMP inhibition were indistinguishable from cells expressing the wild-type μ-opioid receptor stimulated with the agonist DAMGO (right graph, gray bar). Modified from reference [137] with permission.
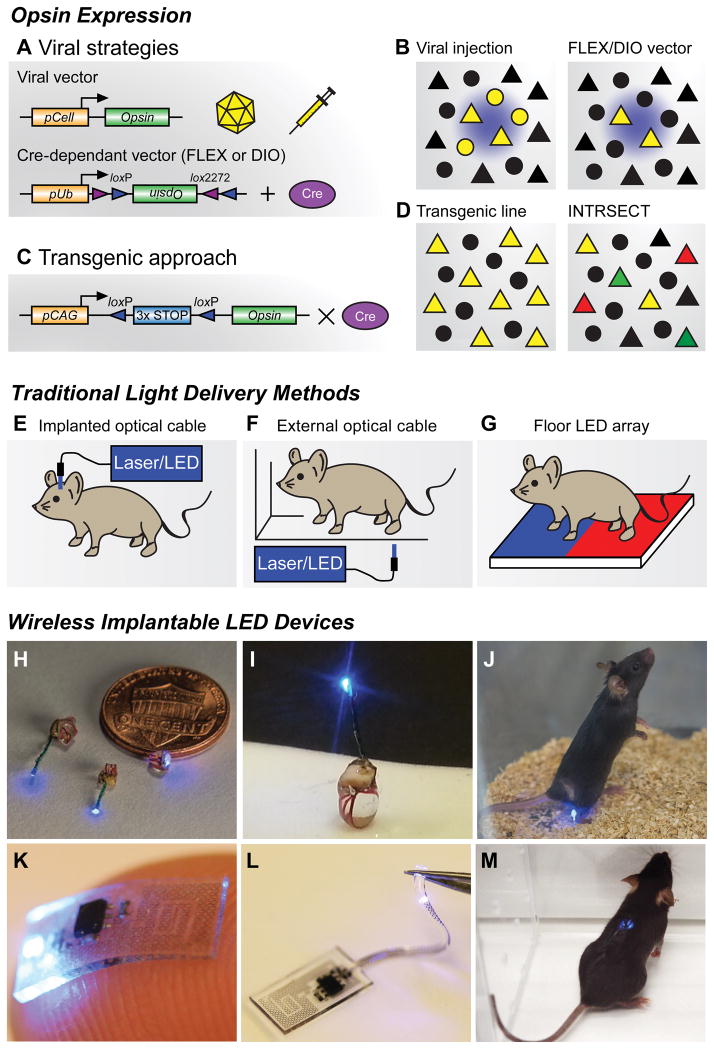
(A) Viral vector strategies for delivering opsins in vivo. Transgene delivery most commonly uses injections of adeno-associated viral (AAV) vectors, though Herpes simplex virus and lentivirus can also be used. Specificity can be achieved by using a promoter of interest to restrict expression to specific cell types (pCell, top), however this approach can lead to weak or non-specific opsin expression. Cre-recombinase dependent vectors using flip-excision (FLEX) or double inverted orientation (DIO) targeting approaches (bottom). An inverted opsin gene (inactive) is flanked with two non-homologous recombination sites (loxP and lox2272). In the presence of Cre recombinase, the opsin is excised and flipped into a functional orientation. Because specificity is generated through tissue-specific expression of Cre-recombinase, strong ubiquitous promoters (pUb) can be used; however, these viruses must be injected in Cre-expressing transgenic animals (Cre database - http://www.gensat.org/cre.jsp ). (B) Cartoon illustrating focal transgene expression (yellow) from a non-Cre-dependent construct in a population of cells near the injection site (left). Similar example illustrating viral delivery of a FLEX/DIO vector into a transgenic animal with Cre-recombinase, depicted here with triangles (right). (C) Transgenic approach for expressing opsins in vivo. Opsin constructs can be introduced into genetic loci to generate transgenic animals for conditional expression. In this approach, opsin genes are preceded by multiple stop codons to prevent expression. These stop codons are flanked by loxP sites, which are excised in the presence of Cre recombinase, generating strong cell-type and region-specific expression. (D) Crossing transgenic opsin lines with Cre driver mice can grant optogenetic access to large neuronal populations (left). Further genetic refinement can be achieved using intersectional genetic approaches like INTRSECT (right). Here cellular specificity is controlled by using multiple recombination events with different enzymes. One group of genetically-defined cells is depicted in green, while a separate population is red. Yellow triangles represent the targeted subpopulation of cells that exhibit genetic features of both of groups. (E) Different brain regions can be anatomically targeted by implanting optical fibers into cannulas affixed to the skull. Light is frequently delivered through a laser or LED light source; however, mice must be tethered to optical cables during behavioral experiments. (F) Optical fibers can be used to illuminate external tissues with light from a laser or LED light source. While animals are untethered, access to deeper tissues is not possible, and uniform light delivery can be problematic. (G) Opsins expressed in peripheral tissues of the paw can be stimulated by LED arrays placed in the floor of behavioral chambers, allowing for untethered movement during experiments. This approach can easily incorporate different stimulation wavelengths. (H,I) Wireless implantable LED devices for stimulating superficial areas in the brain, spinal cord, and peripheral tissues. Reproduced from reference [68] with permission. (J) Image of a freely moving animal with a LED device implanted in the hindpaw. Wireless activation is achieved via radio frequency waves generated by a resonant cavity below the chamber. This couples electromagnetic energy to the mouse, which is harvested by the implant. Reproduced from reference [68] with permission. (K,L) Flexible wireless μLED devices designed to directly interface with the sciatic nerve (K), or be threaded into the epidural space of the spinal column to spinal cord (L) for optogenetic stimulation. Reproduced from reference [122] with permission. (M) Image of a freely moving mouse with a flexible device implanted in the spinal column. Wireless activation is achieved by an external radio frequency antenna that directly powers the μLED device. Reproduced from reference [122] with permission.
Similar articles
-
Optogenetic approaches for investigating neural pathways implicated in schizophrenia and related disorders.Hum Mol Genet. 2014 Sep 15;23(R1):R64-8. doi: 10.1093/hmg/ddu225. Epub 2014 May 13. Hum Mol Genet. 2014. PMID: 24824218 Review.
-
Illuminating neural circuits and behaviour in Caenorhabditis elegans with optogenetics.Philos Trans R Soc Lond B Biol Sci. 2015 Sep 19;370(1677):20140212. doi: 10.1098/rstb.2014.0212. Philos Trans R Soc Lond B Biol Sci. 2015. PMID: 26240427 Free PMC article. Review.
-
Optogenetic inhibition of behavior with anion channelrhodopsins.Nat Methods. 2017 Mar;14(3):271-274. doi: 10.1038/nmeth.4148. Epub 2017 Jan 23. Nat Methods. 2017. PMID: 28114289
-
An excitatory lateral hypothalamic circuit orchestrating pain behaviors in mice.Elife. 2021 May 27;10:e66446. doi: 10.7554/eLife.66446. Elife. 2021. PMID: 34042586 Free PMC article.
-
Functional and Optogenetic Approaches to Discovering Stable Subtype-Specific Circuit Mechanisms in Depression.Biol Psychiatry Cogn Neurosci Neuroimaging. 2019 Jun;4(6):554-566. doi: 10.1016/j.bpsc.2019.04.013. Epub 2019 May 10. Biol Psychiatry Cogn Neurosci Neuroimaging. 2019. PMID: 31176387 Free PMC article.
Cited by
-
In Vivo Interrogation of Spinal Mechanosensory Circuits.Cell Rep. 2016 Nov 1;17(6):1699-1710. doi: 10.1016/j.celrep.2016.10.010. Cell Rep. 2016. PMID: 27806306 Free PMC article.
-
Closed-loop functional optogenetic stimulation.Nat Commun. 2018 Dec 13;9(1):5303. doi: 10.1038/s41467-018-07721-w. Nat Commun. 2018. PMID: 30546051 Free PMC article.
-
Thalamic Bursts and the Epic Pain Model.Front Comput Neurosci. 2017 Jan 12;10:147. doi: 10.3389/fncom.2016.00147. eCollection 2016. Front Comput Neurosci. 2017. PMID: 28127285 Free PMC article. No abstract available.
-
Reticular Formation and Pain: The Past and the Future.Front Neuroanat. 2017 Jul 5;11:51. doi: 10.3389/fnana.2017.00051. eCollection 2017. Front Neuroanat. 2017. PMID: 28725185 Free PMC article. Review.
-
Optogenetic recruitment of spinal reflex pathways from large-diameter primary afferents in non-transgenic rats transduced with AAV9/Channelrhodopsin 2.J Physiol. 2019 Oct;597(19):5025-5040. doi: 10.1113/JP278292. Epub 2019 Aug 28. J Physiol. 2019. PMID: 31397900 Free PMC article.
References
-
- Adamantidis AR, Zhang F, de Lecea L, Deisseroth K. Establishing a fiber-optic-based optical neural interface. Cold Spring Harb Protoc. 2014;2014:839–44. - PubMed
-
- Airan RD, Thompson KR, Fenno LE, Bernstein H, Deisseroth K. Temporally precise in vivo control of intracellular signalling. Nature. 2009;458:1025–1029. - PubMed
-
- Al-Hasani R, McCall JG, Shin G, Gomez AM, Schmitz GP, Bernardi JM, Pyo C-O, Park S, II, Marcinkiewcz CM, Crowley NA, Krashes MJ, Lowell BB, Kash TL, Rogers JA, Bruchas MR. Distinct subpopulations of nucleus accumbens dynorphin neurons drive aversion and reward. Neuron. 2015;87:1063–77. - PMC - PubMed
-
- Aravanis AM, Wang L-P, Zhang F, Meltzer La, Mogri MZ, Schneider MB, Deisseroth K. An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. J Neural Eng. 2007;4:S143–S156. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
