

β2-Adrenergic Receptors Chaperone Trapped Bitter Taste Receptor 14 to the Cell Surface as a Heterodimer and Exert Unidirectional Desensitization of Taste Receptor Function
- PMID: 27342779
- PMCID: PMC5016158
- DOI: 10.1074/jbc.M116.722736
β2-Adrenergic Receptors Chaperone Trapped Bitter Taste Receptor 14 to the Cell Surface as a Heterodimer and Exert Unidirectional Desensitization of Taste Receptor Function
Abstract
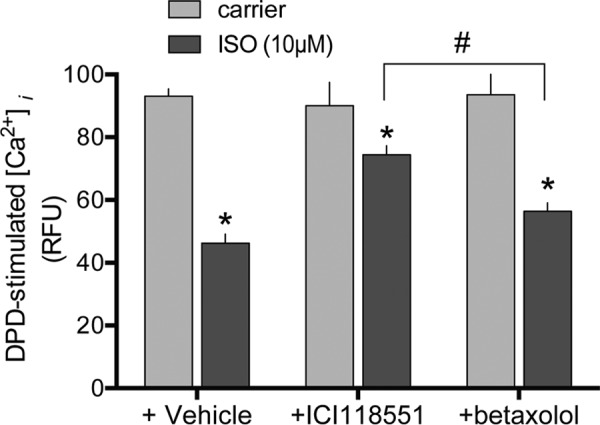
Bitter taste receptors (TAS2Rs) are G-protein-coupled receptors now recognized to be expressed on extraoral cells, including airway smooth muscle (ASM) where they evoke relaxation. TAS2Rs are difficult to express in heterologous systems, with most receptors being trapped intracellularly. We find, however, that co-expression of β2-adrenergic receptors (β2AR) in HEK-293T routes TAS2R14 to the cell surface by forming receptor heterodimers. Cell surface TAS2R14 expression was increased by ∼5-fold when β2AR was co-expressed. Heterodimer formation was shown by co-immunoprecipitation with tagged receptors, biomolecular fluorescence complementation, and merged confocal images. The dynamic nature of this interaction was shown by: a gene-dose relationship between transfected β2AR and TAS2R14 expression, enhanced (up to 3-fold) TAS2R14 agonist stimulation of [Ca(2+)]i with β2AR co-transfection, ∼53% decrease in [Ca(2+)]i signaling with shRNA knockdown of β2AR in H292 cells, and ∼60% loss of [Ca(2+)]i responsiveness in βAR knock-out mouse ASM. Once expressed on the surface, we detected unidirectional, conformation-dependent, interaction within the heterodimer, with β2AR activation rapidly uncoupling TAS2R14 function (∼65% desensitization). Cross-talk was independent of β2AR internalization and cAMP/PKA, and not accompanied by TAS2R14 internalization. With prolonged β-agonist exposure, TAS2R14 internalized, consistent with slow recycling of naked TAS2R14 in the absence of the heterodimeric milieu. In studies of ASM mechanics, rapid cross-talk was confirmed at the physiologic level, where relaxation from TAS2R14 agonist was decreased by ∼50% with β-agonist co-treatment. Thus the β2AR acts as a double-edged sword: increasing TAS2R14 cell surface expression, but when activated by β-agonist, partially offsetting the expression phenotype by direct receptor:receptor desensitization of TAS2R14 function.
Keywords: G protein; asthma; bitter taste receptor; chronic obstructive pulmonary disease (COPD); cyclic AMP (cAMP); dimerization; receptor internalization; smooth muscle.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures










References
-
- Chandrashekar J., Hoon M. A., Ryba N. J., and Zuker C. S. (2006) The receptors and cells for mammalian taste. Nature 444, 288–294 - PubMed
-
- Meyerhof W., Batram C., Kuhn C., Brockhoff A., Chudoba E., Bufe B., Appendino G., and Behrens M. (2010) The molecular receptive ranges of human TAS2R bitter taste receptors. Chem. Senses 35, 157–170 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
