

Negative feedback regulation of the ERK1/2 MAPK pathway
- PMID: 27342992
- PMCID: PMC5075022
- DOI: 10.1007/s00018-016-2297-8
Negative feedback regulation of the ERK1/2 MAPK pathway
Abstract
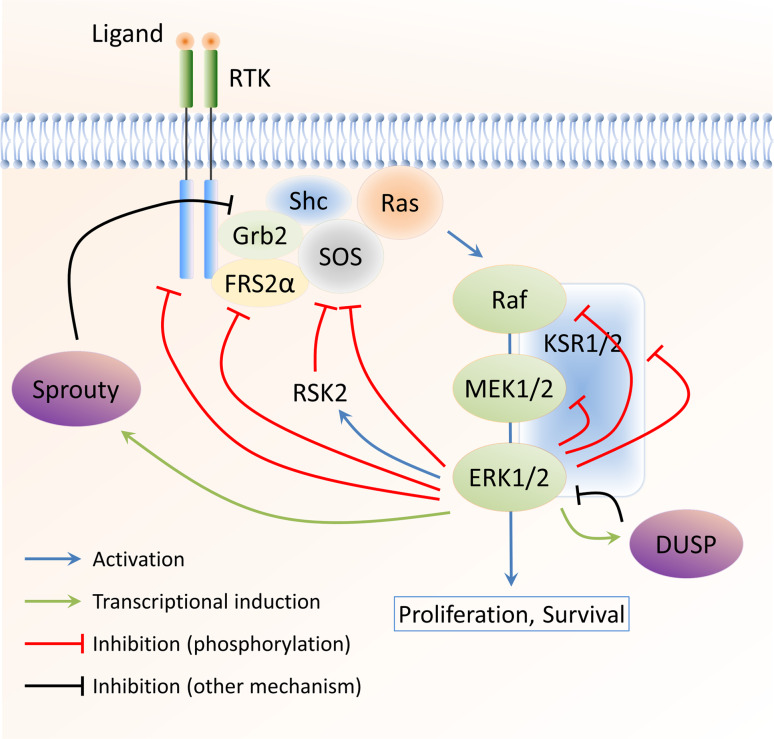
The extracellular signal-regulated kinase 1/2 (ERK1/2) mitogen-activated protein kinase (MAPK) signalling pathway regulates many cellular functions, including proliferation, differentiation, and transformation. To reliably convert external stimuli into specific cellular responses and to adapt to environmental circumstances, the pathway must be integrated into the overall signalling activity of the cell. Multiple mechanisms have evolved to perform this role. In this review, we will focus on negative feedback mechanisms and examine how they shape ERK1/2 MAPK signalling. We will first discuss the extensive number of negative feedback loops targeting the different components of the ERK1/2 MAPK cascade, specifically the direct posttranslational modification of pathway components by downstream protein kinases and the induction of de novo gene synthesis of specific pathway inhibitors. We will then evaluate how negative feedback modulates the spatiotemporal signalling dynamics of the ERK1/2 pathway regarding signalling amplitude and duration as well as subcellular localisation. Aberrant ERK1/2 activation results in deregulated proliferation and malignant transformation in model systems and is commonly observed in human tumours. Inhibition of the ERK1/2 pathway thus represents an attractive target for the treatment of malignant tumours with increased ERK1/2 activity. We will, therefore, discuss the effect of ERK1/2 MAPK feedback regulation on cancer treatment and how it contributes to reduced clinical efficacy of therapeutic agents and the development of drug resistance.
Keywords: Cancer; Cell signalling; Negative feedback; Pathway modelling; Signalling dynamics; Spatiotemporal regulation.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
