

Two types of soybean diacylglycerol acyltransferases are differentially involved in triacylglycerol biosynthesis and response to environmental stresses and hormones
- PMID: 27345221
- PMCID: PMC4921965
- DOI: 10.1038/srep28541
Two types of soybean diacylglycerol acyltransferases are differentially involved in triacylglycerol biosynthesis and response to environmental stresses and hormones
Abstract
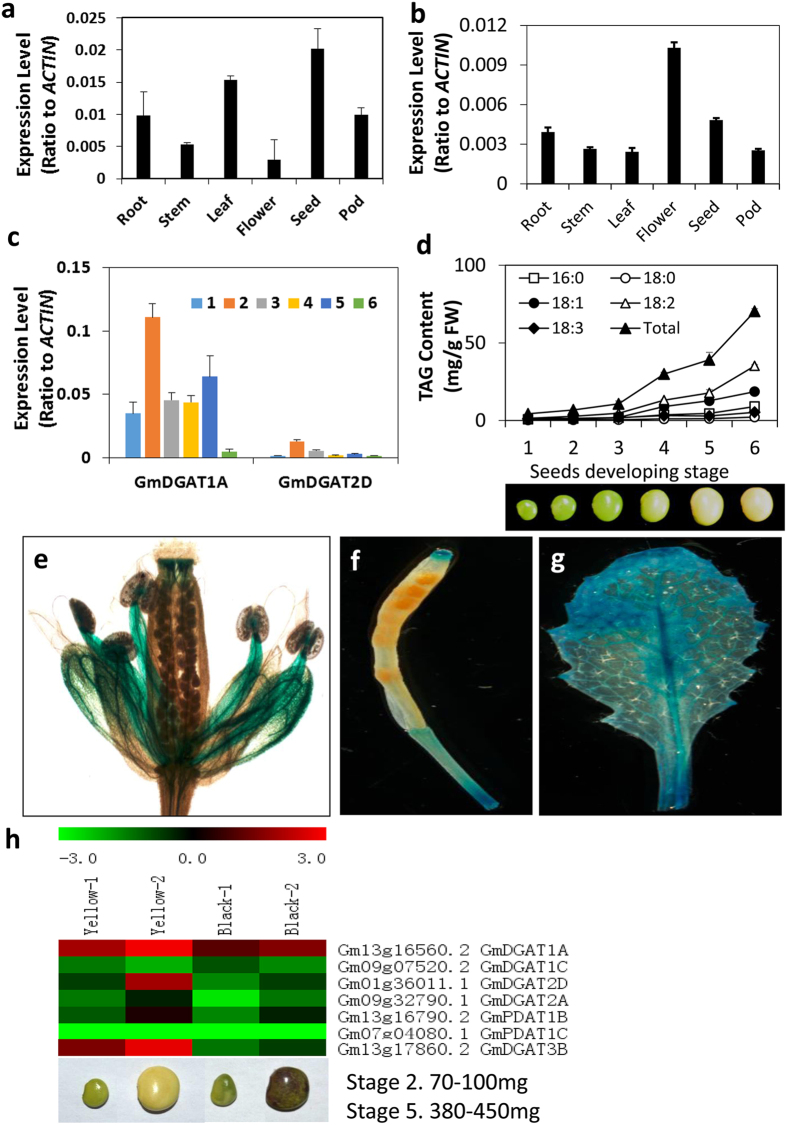
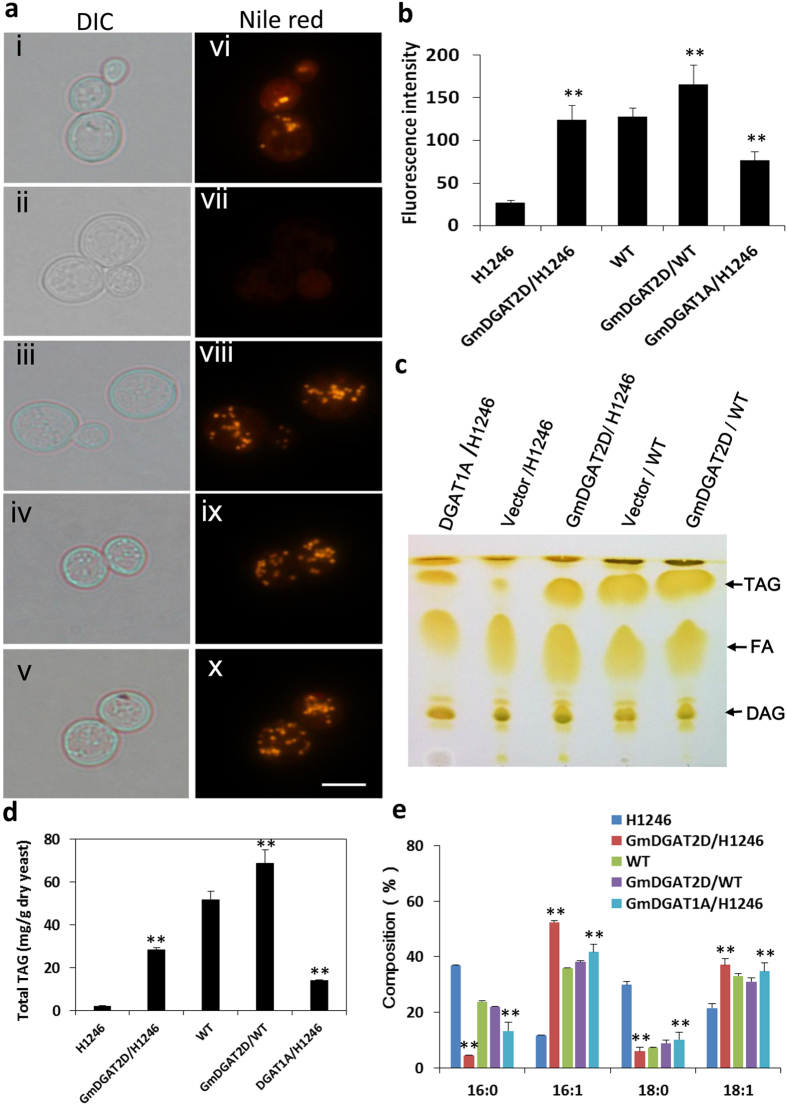
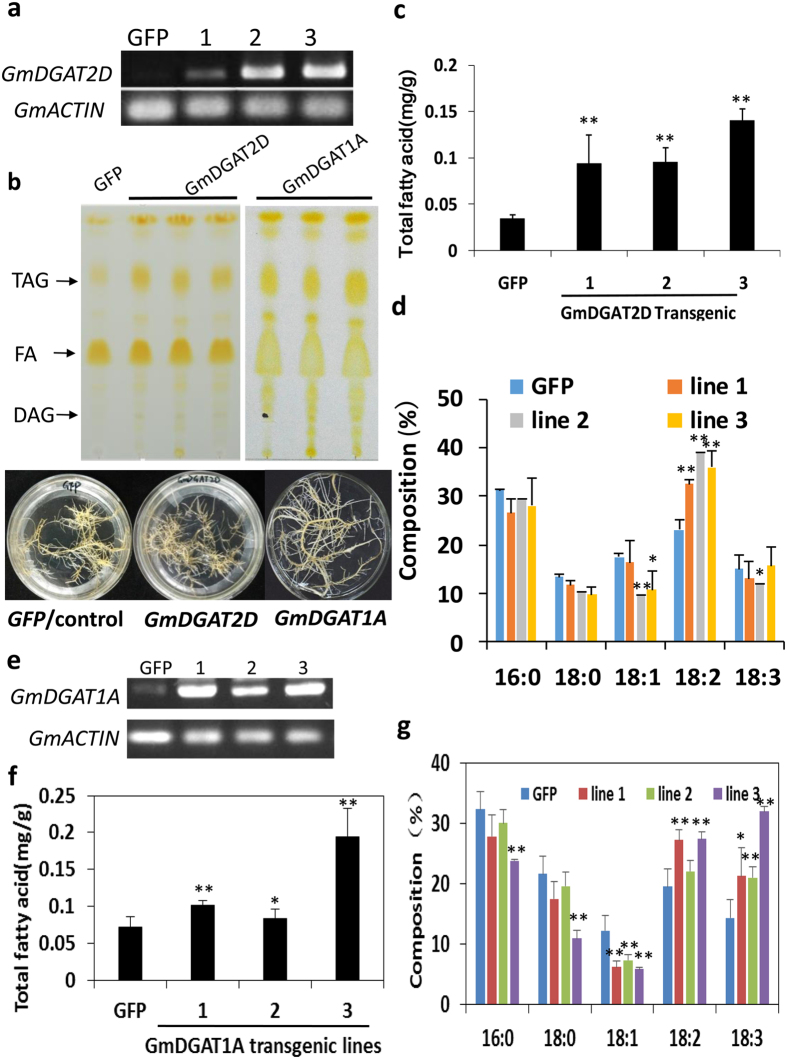
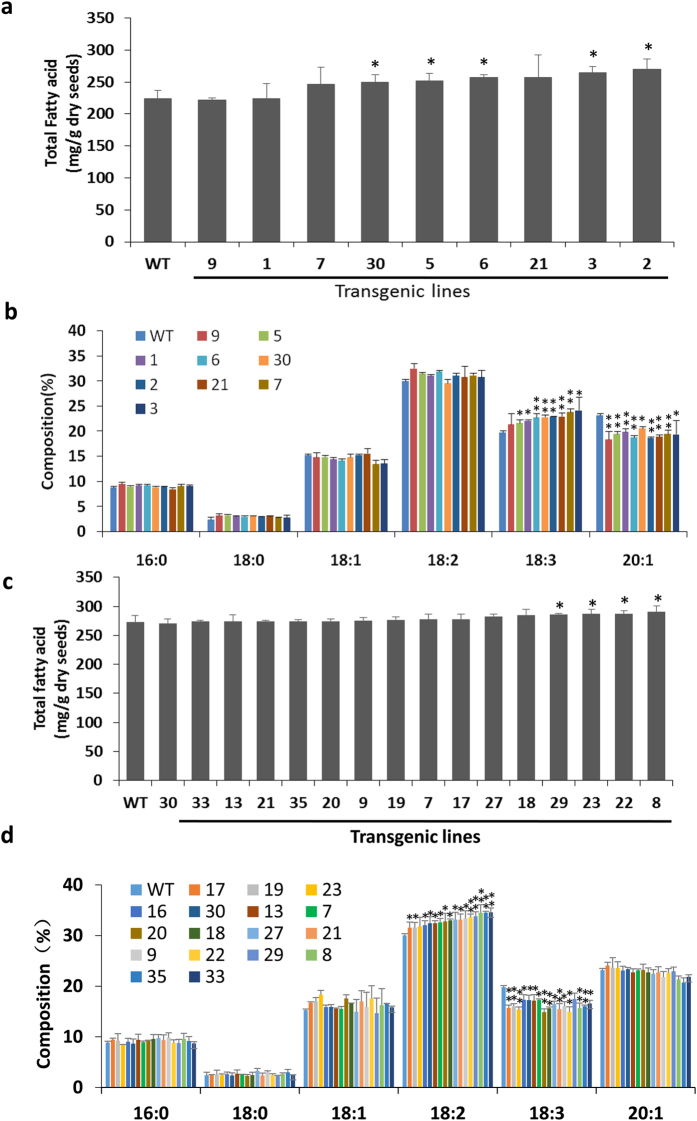
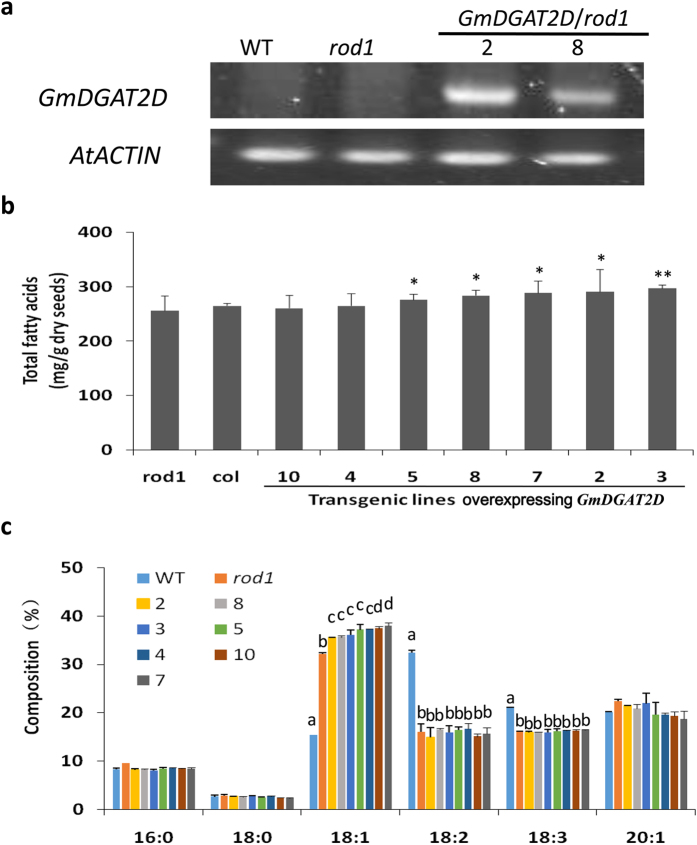
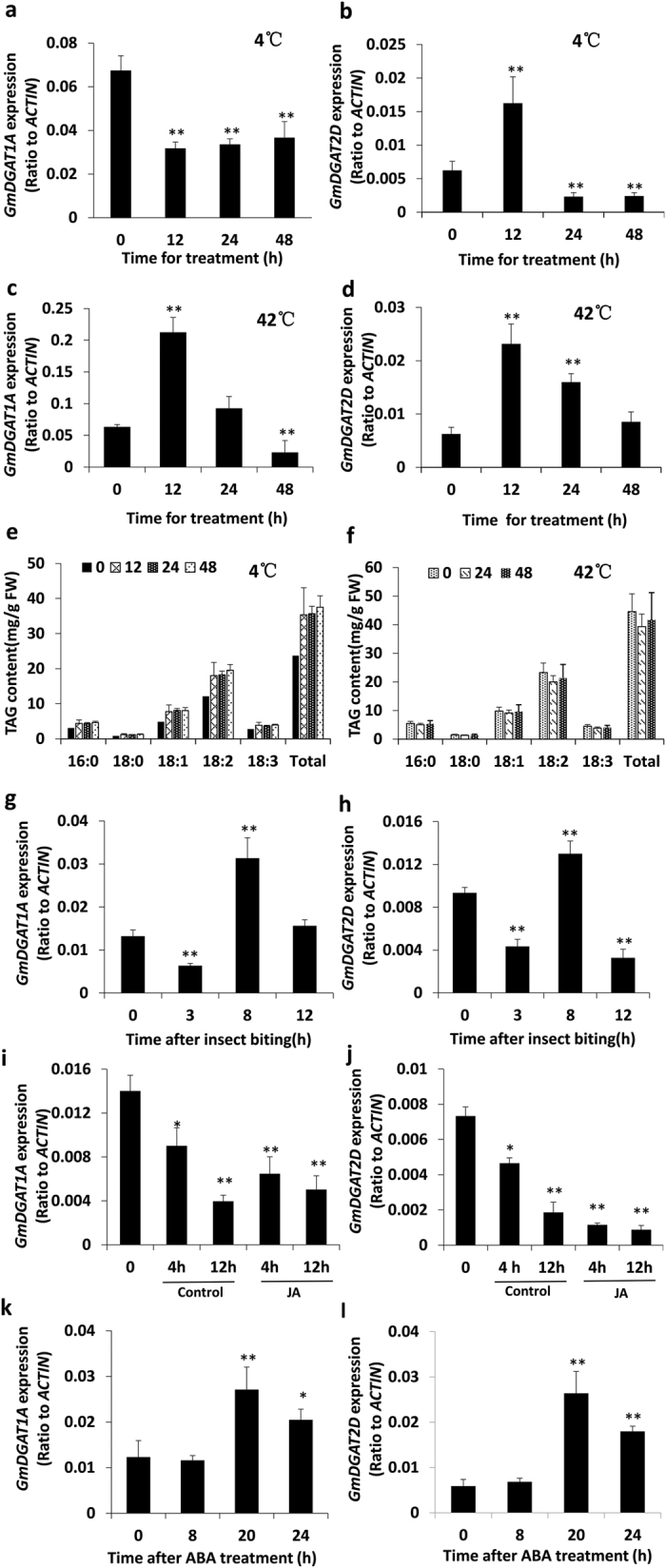
Diacylglycerol acyltransferases (DGATs) play a key role in plant triacylglycerol (TAG) biosynthesis. Two type 1 and 2 DGATs from soybean were characterized for their functions in TAG biosynthesis and physiological roles. GmDGAT1A is highly expressed in seeds while GmDGAT2D is mainly expressed in flower tissues. They showed different expression patterns in response to biotic and abiotic stresses. GmDGAT2D was up-regulated by cold and heat stress and ABA signaling, and repressed by insect biting and jasmonate, whereas GmDGAT1A show fewer responses. Both GmDGAT1A and GmDGAT2D were localized to the endoplasmic reticulum and complemented the TAG deficiency of a yeast mutant H1246. GmDGAT2D-transgenic hairy roots synthesized more 18:2- or 18:1-TAG, whereas GmDGAT1A prefers to use 18:3-acyl CoA for TAG synthesis. Overexpression of both GmDGATs in Arabidopsis seeds enhanced the TAG production; GmDGAT2D promoted 18:2-TAG in wild-type but enhanced 18:1-TAG production in rod1 mutant seeds, with a decreased 18:3-TAG. However, GmDGAT1A enhanced 18:3-TAG and reduced 20:1-TAG contents. The different substrate preferences of two DGATs may confer diverse fatty acid profiles in soybean oils. While GmDGAT1A may play a role in usual seed TAG production and GmDGAT2D is also involved in usual TAG biosynthesis in other tissues in responses to environmental and hormonal cues.
Figures

References
-
- Wilson R. F. The role of genomics and biotechnology in achieving global food security for high-oleic vegetable oil. Journal of oleo science 61, 357–367 (2012). - PubMed
-
- Singer S. D., Zou J. & Weselake R. J. Abiotic factors influence plant storage lipid accumulation and composition. Plant Sci 243, 1–9 (2016). - PubMed
-
- Moellering E. R., Muthan B. & Benning C. Freezing tolerance in plants requires lipid remodeling at the outer chloroplast membrane. Science 330, 226–228 (2000). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
