

Molecular basis for high affinity and selectivity of peptide antagonist, Bantag-1, for the orphan BB3 receptor
- PMID: 27346274
- PMCID: PMC4980153
- DOI: 10.1016/j.bcp.2016.06.013
Molecular basis for high affinity and selectivity of peptide antagonist, Bantag-1, for the orphan BB3 receptor
Abstract
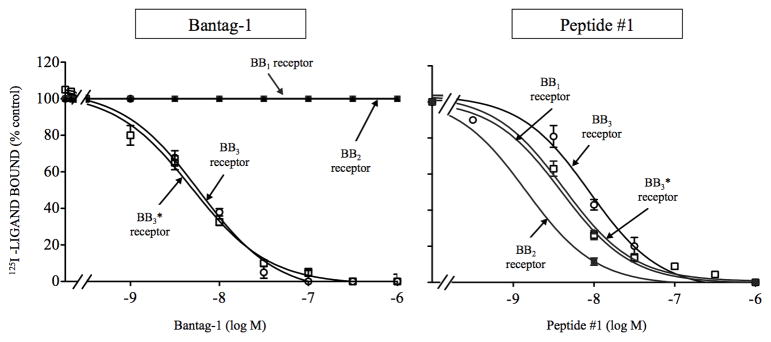
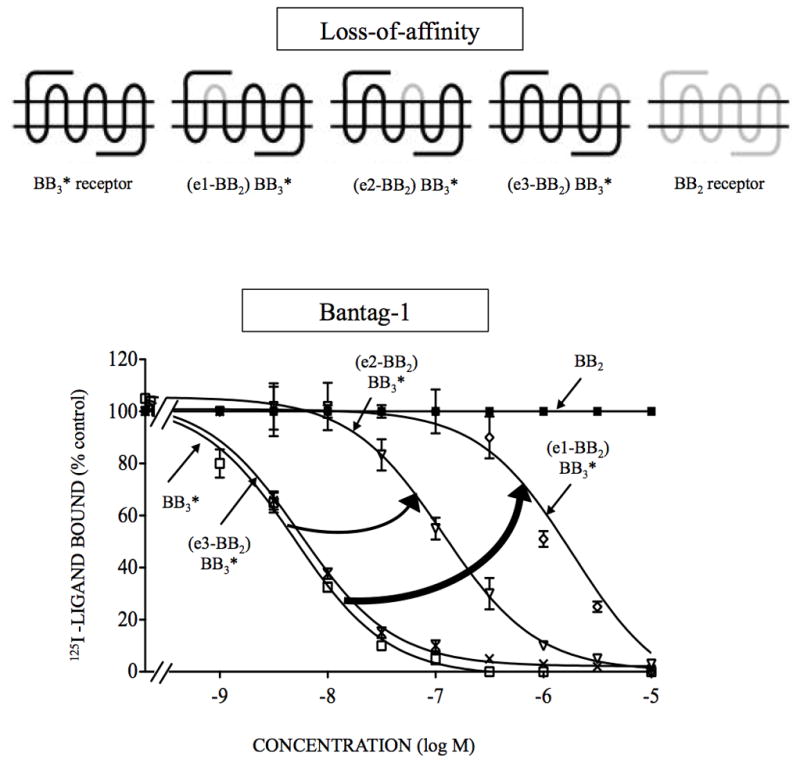
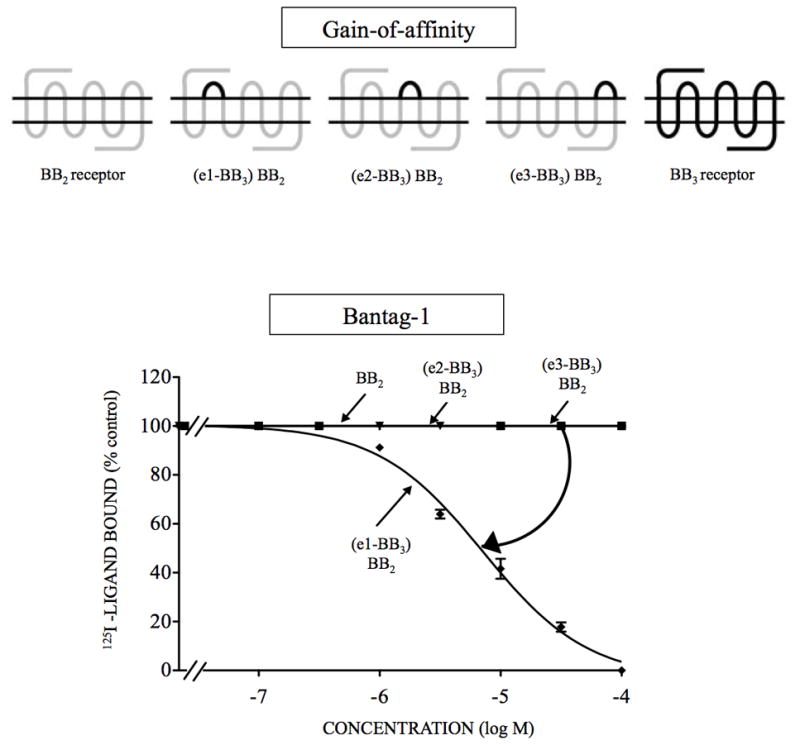
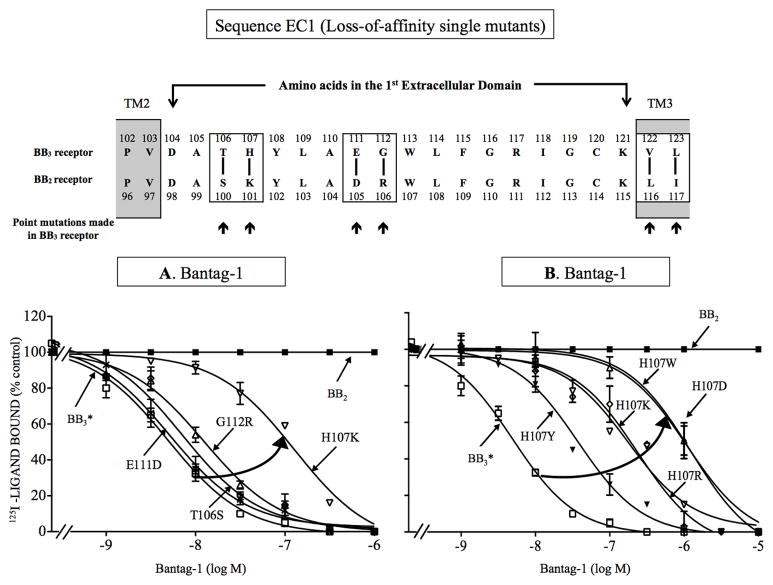
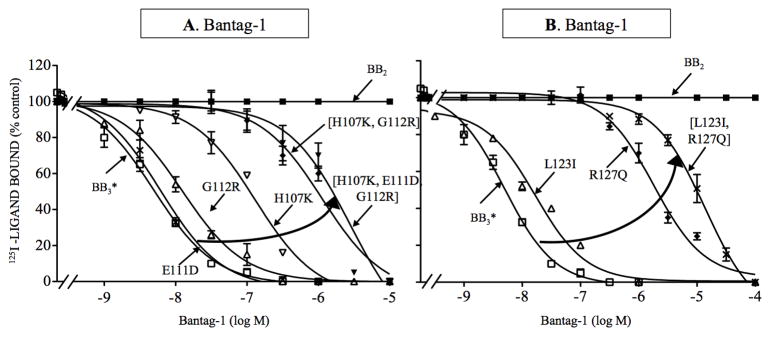
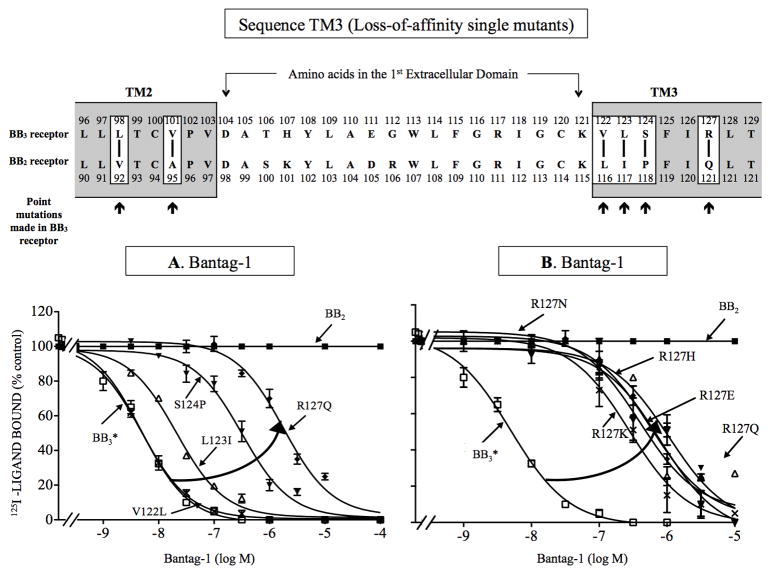
Bombesin-receptor-subtype-3 (BB3 receptor) is a G-protein-coupled-orphan-receptor classified in the mammalian Bombesin-family because of high homology to gastrin-releasing peptide (BB2 receptor)/neuromedin-B receptors (BB1 receptor). There is increased interest in BB3 receptor because studies primarily from knockout-mice suggest it plays roles in energy/glucose metabolism, insulin-secretion, as well as motility and tumor-growth. Investigations into its roles in physiological/pathophysiological processes are limited because of lack of selective ligands. Recently, a selective, peptide-antagonist, Bantag-1, was described. However, because BB3 receptor has low-affinity for all natural, Bn-related peptides, little is known of the molecular basis of its high-affinity/selectivity. This was systematically investigated in this study for Bantag-1 using a chimeric-approach making both Bantag-1 loss-/gain-of-affinity-chimeras, by exchanging extracellular (EC) domains of BB3/BB2 receptor, and using site-directed-mutagenesis. Receptors were transiently expressed and affinities determined by binding studies. Bantag-1 had >5000-fold selectivity for BB3 receptor over BB2/BB1 receptors and substitution of the first EC-domain (EC1) in loss-/gain-of affinity-chimeras greatly affected affinity. Mutagenesis of each amino acid difference in EC1 between BB3 receptor/BB2 receptor showed replacement of His(107) in BB3 receptor by Lys(107) (H107K-BB3 receptor-mutant) from BB2 receptor, decreased affinity 60-fold, and three replacements [H107K, E11D, G112R] decreased affinity 500-fold. Mutagenesis in EC1's surrounding transmembrane-regions (TMs) demonstrated TM2 differences were not important, but R127Q in TM3 alone decreased affinity 400-fold. Additional mutants in EC1/TM3 explored the molecular basis for these changes demonstrated in EC1, particularly important is the presence of aromatic-interactions by His(107), rather than hydrogen-bonding or charge-charge interactions, for determining Bantag-1 high affinity/selectivity. In regard to Arg(127) in TM3, both hydrogen-bonding and charge-charge interactions contribute to the high-affinity/selectivity for Bantag-1.
Keywords: Bombesin; Gastrin-releasing peptide; Neuromedin B; Obesity; Satiety.
Published by Elsevier Inc.
Conflict of interest statement
The authors have no conflicts of interest with this study.
Figures
Similar articles
-
Molecular basis for agonist selectivity and activation of the orphan bombesin receptor subtype 3 receptor.J Pharmacol Exp Ther. 2008 Feb;324(2):463-74. doi: 10.1124/jpet.107.132332. Epub 2007 Nov 15. J Pharmacol Exp Ther. 2008. PMID: 18006692 Free PMC article.
-
Comparative pharmacology of bombesin receptor subtype-3, nonpeptide agonist MK-5046, a universal peptide agonist, and peptide antagonist Bantag-1 for human bombesin receptors.J Pharmacol Exp Ther. 2013 Oct;347(1):100-16. doi: 10.1124/jpet.113.206896. Epub 2013 Jul 26. J Pharmacol Exp Ther. 2013. PMID: 23892571 Free PMC article.
-
Molecular basis for selectivity of high affinity peptide antagonists for the gastrin-releasing peptide receptor.J Biol Chem. 2001 Sep 28;276(39):36652-63. doi: 10.1074/jbc.M104566200. Epub 2001 Aug 2. J Biol Chem. 2001. PMID: 11463790
-
99mTc-N2S2-Tat(49-57)-Lys3-bombesin.2009 Aug 10 [updated 2009 Sep 23]. In: Molecular Imaging and Contrast Agent Database (MICAD) [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2004–2013. 2009 Aug 10 [updated 2009 Sep 23]. In: Molecular Imaging and Contrast Agent Database (MICAD) [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2004–2013. PMID: 20641937 Free Books & Documents. Review.
-
99mTc(v)O-Gly-Gly-Cys-Orn-Orn-Orn-Bombesin[2-14].2009 Aug 19 [updated 2009 Sep 23]. In: Molecular Imaging and Contrast Agent Database (MICAD) [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2004–2013. 2009 Aug 19 [updated 2009 Sep 23]. In: Molecular Imaging and Contrast Agent Database (MICAD) [Internet]. Bethesda (MD): National Center for Biotechnology Information (US); 2004–2013. PMID: 20641903 Free Books & Documents. Review.
Cited by
-
Development and Characterization of a Novel, High-Affinity, Specific, Radiolabeled Ligand for BRS-3 Receptors.J Pharmacol Exp Ther. 2019 Jun;369(3):454-465. doi: 10.1124/jpet.118.255141. Epub 2019 Apr 10. J Pharmacol Exp Ther. 2019. PMID: 30971479 Free PMC article.
-
AM-37 and ST-36 Are Small Molecule Bombesin Receptor Antagonists.Front Endocrinol (Lausanne). 2017 Jul 21;8:176. doi: 10.3389/fendo.2017.00176. eCollection 2017. Front Endocrinol (Lausanne). 2017. PMID: 28785244 Free PMC article.
-
Understanding Peptide Binding in Class A G Protein-Coupled Receptors.Mol Pharmacol. 2019 Nov;96(5):550-561. doi: 10.1124/mol.119.115915. Epub 2019 Jul 10. Mol Pharmacol. 2019. PMID: 31436539 Free PMC article. Review.
-
Neuropeptide G Protein-Coupled Receptors as Oncotargets.Front Endocrinol (Lausanne). 2018 Jun 29;9:345. doi: 10.3389/fendo.2018.00345. eCollection 2018. Front Endocrinol (Lausanne). 2018. PMID: 30008698 Free PMC article. Review.
-
The Nonpeptide Agonist MK-5046 Functions As an Allosteric Agonist for the Bombesin Receptor Subtype-3.J Pharmacol Exp Ther. 2022 Aug;382(2):66-78. doi: 10.1124/jpet.121.001033. Epub 2022 May 29. J Pharmacol Exp Ther. 2022. PMID: 35644465 Free PMC article.
References
-
- Fathi Z, Corjay MH, Shapira H, Wada E, Benya R, Jensen R, et al. BRS-3: novel bombesin receptor subtype selectively expressed in testis and lung carcinoma cells. J Biol Chem. 1993;268(8):5979–5984. - PubMed
-
- Majumdar ID, Weber HC. Biology and pharmacology of bombesin receptor subtype-3. Curr Opin Endocrinol Diabetes Obes. 2012;19:3–7. - PubMed
-
- Majumdar ID, Weber HC. Biology of mammalian bombesin-like peptides and their receptors. Curr Opin Endocrinol Diabetes Obes. 2011;18:68–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
