

5-Hydroxymethylcytosine Remodeling Precedes Lineage Specification during Differentiation of Human CD4(+) T Cells
- PMID: 27346350
- PMCID: PMC5868728
- DOI: 10.1016/j.celrep.2016.05.091
5-Hydroxymethylcytosine Remodeling Precedes Lineage Specification during Differentiation of Human CD4(+) T Cells
Abstract
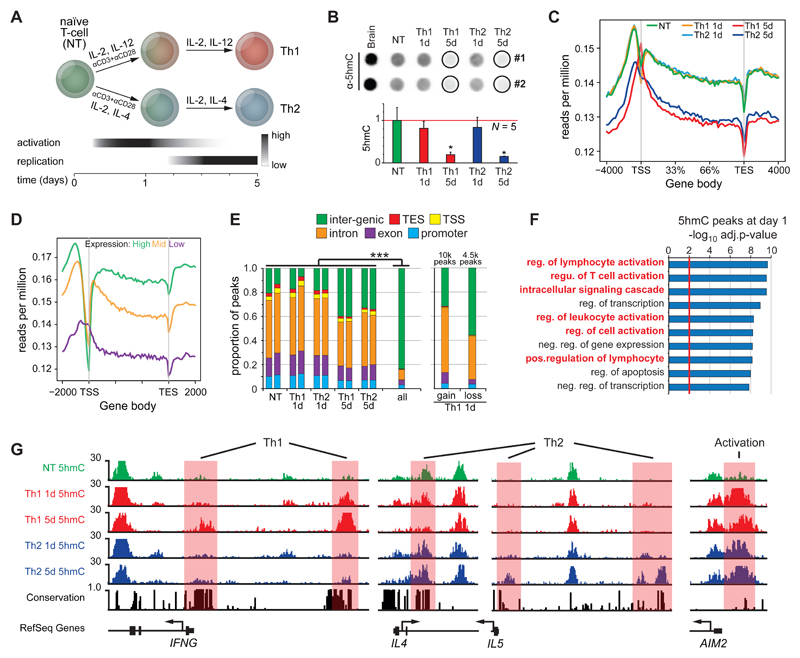
5-methylcytosine (5mC) is converted to 5-hydroxymethylcytosine (5hmC) by the TET family of enzymes as part of a recently discovered active DNA de-methylation pathway. 5hmC plays important roles in regulation of gene expression and differentiation and has been implicated in T cell malignancies and autoimmunity. Here, we report early and widespread 5mC/5hmC remodeling during human CD4(+) T cell differentiation ex vivo at genes and cell-specific enhancers with known T cell function. We observe similar DNA de-methylation in CD4(+) memory T cells in vivo, indicating that early remodeling events persist long term in differentiated cells. Underscoring their important function, 5hmC loci were highly enriched for genetic variants associated with T cell diseases and T-cell-specific chromosomal interactions. Extensive functional validation of 22 risk variants revealed potentially pathogenic mechanisms in diabetes and multiple sclerosis. Our results support 5hmC-mediated DNA de-methylation as a key component of CD4(+) T cell biology in humans, with important implications for gene regulation and lineage commitment.
Copyright © 2016 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
