

doi: 10.1038/srep28107.
Identifying Kinase Substrates via a Heavy ATP Kinase Assay and Quantitative Mass Spectrometry
Affiliations
- PMID: 27346722
- PMCID: PMC4921819
- DOI: 10.1038/srep28107
Item in Clipboard
Identifying Kinase Substrates via a Heavy ATP Kinase Assay and Quantitative Mass Spectrometry
Sci Rep.
.
Abstract
Mass spectrometry-based in vitro kinase screens play an essential role in the discovery of kinase substrates, however, many suffer from biological and technical noise or necessitate genetically-altered enzyme-cofactor systems. We describe a method that combines stable γ-[(18)O2]-ATP with classical in vitro kinase assays within a contemporary quantitative proteomic workflow. Our approach improved detection of known substrates of the non-receptor tyrosine kinase ABL1; and identified potential, new in vitro substrates.
Figures
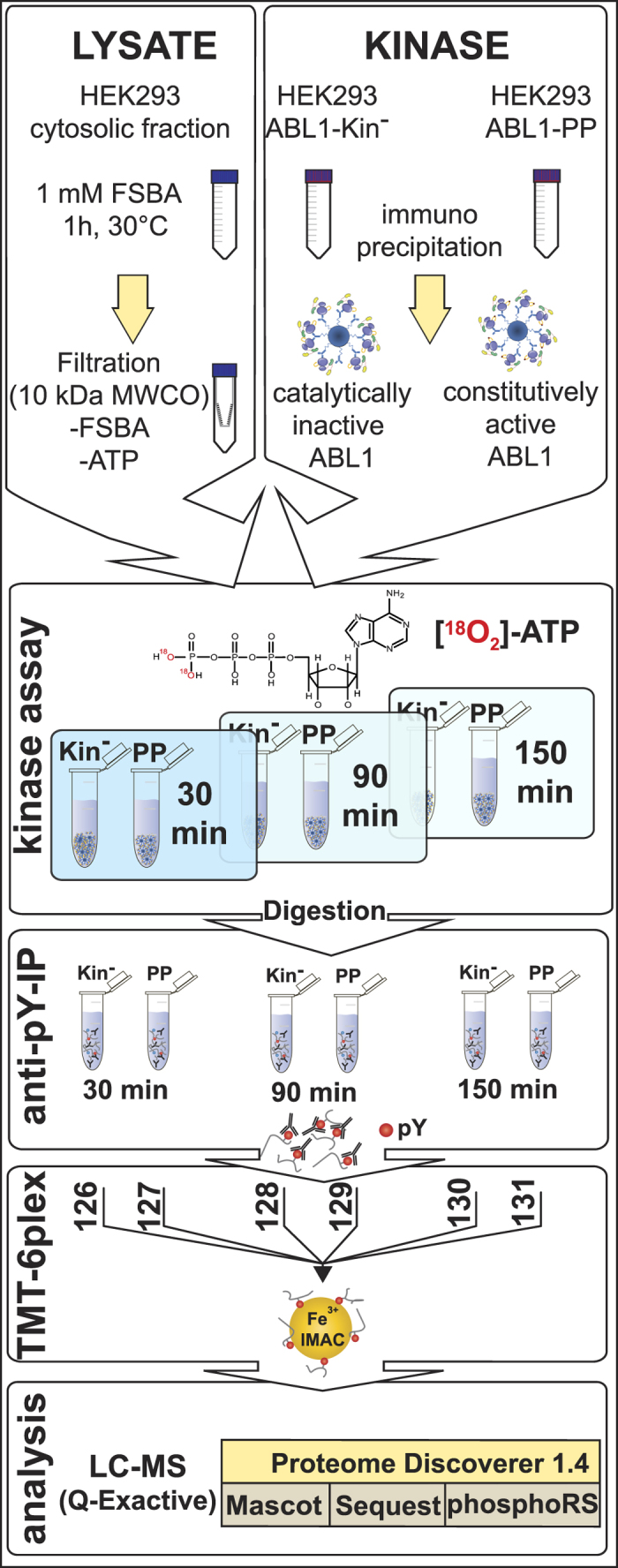
Cytosolic fractions were treated with the pan-kinase inhibitor FSBA to irreversibly abolish cellular kinase activity. Simultaneous exchange to the kinase assay buffer; and removal of excess FSBA and endogenous ATP was achieved by ultrafiltration. In parallel, constitutively-active ABL1-PP and inactive ABL1-Kin− were immunopurified via protein G sepharose beads. Prior to the kinase assay, treated lysate and washed kinase-bead fractions were equally divided, mixed and the kinase reaction initiated with ‘heavy’ 18O-labelled ATP. To obtain the phosphorylation kinetic series, pairs of active/inactive kinase reactions were quenched at defined time points. Following enzymatic digestion, peptides were concentrated with solid-phase extraction and phosphotyrosine-containing peptides enriched via immunoprecipitation. Eluted peptides were chemically-modified with isobaric amine-reactive tandem mass tag (TMT) reagents to multiplex 3 × 2 samples. After a second phosphopeptide enrichment with immobilised metal-affinity chromatography (IMAC), the sample was analysed by LCMS.
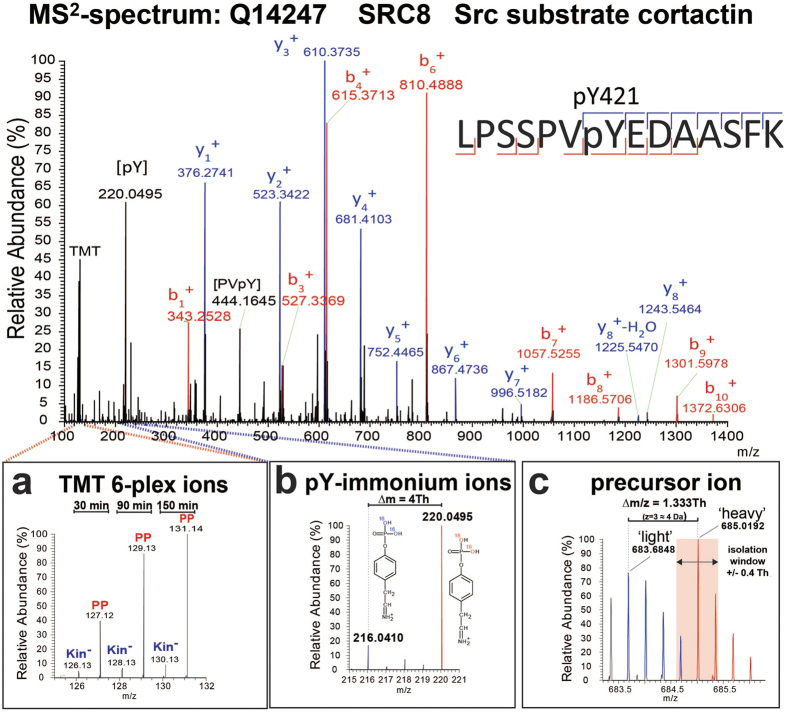
An example of an MS2 spectrum corresponding to a tryptic peptide derived from SRC8 containing a reported ABL1 phosphorylation site. The assigned fragment ions provide peptide-specific sequence information that is used to computationally match the data to the corresponding peptide in a protein database. Certain fragment ions (i.e., y8 and b7) unambiguously assign the phosphate group (+84 Da) to Y421 and not to S417, S418 or S426. (a) Magnification of the TMT reporter ion region to illustrate the relative abundance of the individual multiplexed samples for this specific phosphotyrosine-containing peptide. TMT 6-plex ion intensities were used to generate individual relative ratios or to plot reaction progress kinetics. (b) pY-immonium ions for ‘light’ (m/z 216.04) and ‘heavy’ (m/z 220.04) versions of the phosphopeptide. The existence and relative abundances of the immonium ions provide unequivocal evidence of tyrosine residue phosphorylation and quality control of the respective spectrum. (c) MS1 precursor ion of the phosphotyrosine-containing peptide with both the ‘light’ (m/z 683.6848) and ‘heavy’ (m/z 685.0192) isotopologues. Both isotopologues (blue and red peaks) display a typical 12C/13C-isotopic peak distribution (1 a.m.u. separation) but with a slight overlap due to the small ∆m/z of 1.333 Th. Precursor ion selection of the monoisotopic peak (pink area) was within the isolation window of ±0.4 Th and partially including the ‘light’ isotopologue (fourth 13C-isotopic peak).
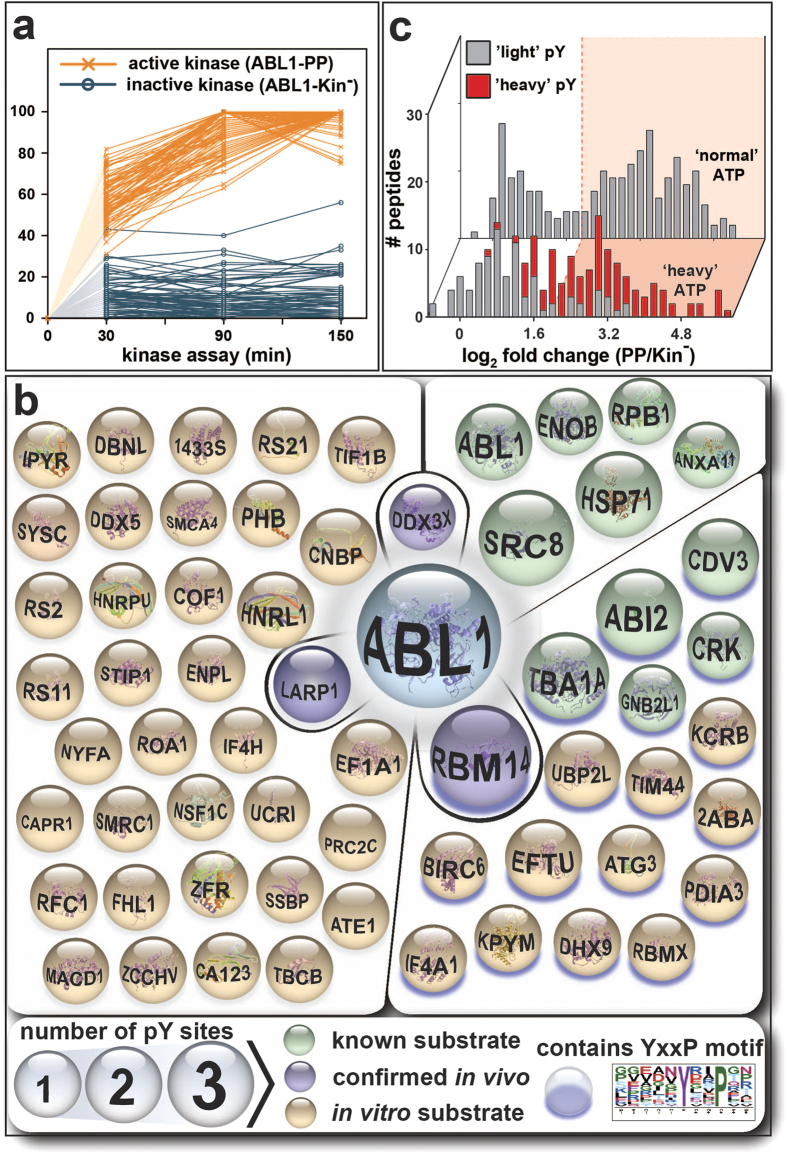
(a) TMT reporter ion intensities of phosphotyrosine-containing peptides were normalised and plotted against the respective kinase assay time point. Connecting lines are shown for active ABL1-PP (orange) or inactive ABL1-Kin− (blue). As ‘heavy’ phosphate groups are absent prior to the addition of 18O-labelled ATP, the lines were extrapolated through the origin (shaded to indicate hypothetical). A clear separation between the kinetic trend lines for active and inactive ABL1 was apparent. (b) All 61 identified in vitro substrates of ABL1 with at least one ‘heavy’ phosphotyrosine residue. Known and validated substrates are shown in green and purple, respectively. Proteins with purple shading contain the preferred ABL1 YxxP consensus motif (p-value ≤0.05; motif-score: 4.63; fold-increase: 3.16; www.motif-x.med.harvard.edu ). Sphere size indicates the number of phosphotyrosines per substrate. (c) Comparison of the ABL1-PP/ABL1-Kin− ratio distribution at 90 minutes for HAKA-MS (front) versus the kinase assay performed with normal ATP (back). ‘Light’ and ‘heavy’ pY-containing peptides are labelled in grey and red, respectively. In both cases, the global distribution is bimodal with a shallow, intersecting valley. For the normal ATP kinase assay, the threshold for putative in vitro ABL1 substrates was approximated with mixed Gaussian clustering. The first and second clusters are considered noise and in vitro substrates, respectively. Comparison to HAKA-MS revealed that regardless of the cut-off stringency (i.e., pink, ≥ 4 ratio), data from the normal ATP assay can contain many false positives (grey). Conversely, true positives (red) were excluded.
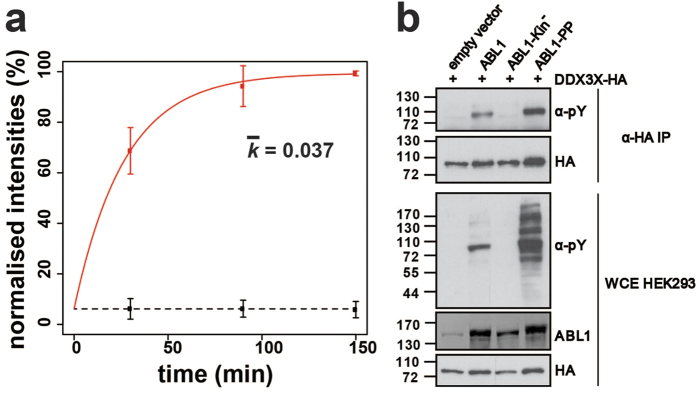
(a) Phosphorylation kinetics for the DDX3X peptide DKDAYSSFGSR. The ‘heavy’ phosphate group is located on Y69. Calculated rate constants (k) are plotted for the constitutively-active ABL1-PP and catalytically-inactive ABL1-Kin−. Error bars represent two standard deviations (2σ) from the mean. (b) Biochemical validation of DDX3X tyrosine phosphorylation mediated by ABL1. HA-tagged DDX3X was transiently co-transfected in HEK293 cells in the presence of ABL1, ABL1-Kin−, ABL1-PP or the empty vector. DDX3X was immunopurified with HA-beads and the tyrosine phosphorylation state assessed by α-phosphotyrosine (4G10) immunostaining. Total levels of DDX3X-HA, ABL1 and tyrosine-phosphorylated proteins were visualised by immunostaining against α-HA, α-ABL1 and α-phosphotyrosine (4G10), respectively.
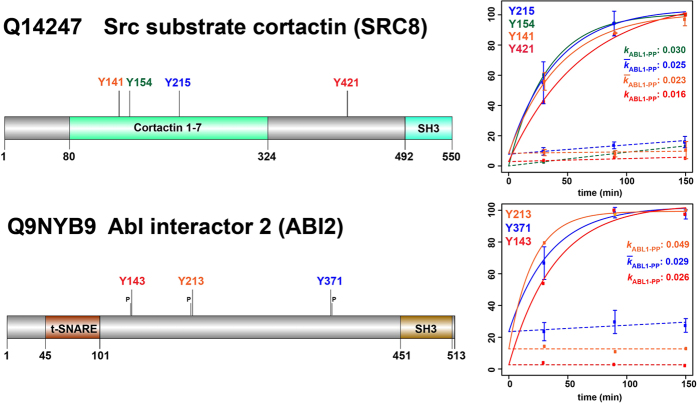
For Src substrate cortactin (SRC8) and ABL interactor 2 (ABI2), multiple ‘heavy’ pY-sites were identified; with the latter containing the YxxP ABL1 consensus motif at all three sites. The plots show TMT 6-plex-based reaction progress kinetics with different rate constants determined for each of the pY-sites.  and
and  , represent rate constant and mean rate constant, respectively. Error bars represent two standard deviations (2σ) from the mean.
, represent rate constant and mean rate constant, respectively. Error bars represent two standard deviations (2σ) from the mean.
and , represent rate constant and mean rate constant, respectively. Error bars represent two standard deviations (2σ) from the mean.References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
