

Review
doi: 10.1038/nri.2016.62.
Epub 2016 Jun 27.
The unfolded protein response in immunity and inflammation
Affiliations
- PMID: 27346803
- PMCID: PMC5310224
- DOI: 10.1038/nri.2016.62
Item in Clipboard
Review
The unfolded protein response in immunity and inflammation
Nat Rev Immunol.
2016 Aug.
Abstract
The unfolded protein response (UPR) is a highly conserved pathway that allows the cell to manage endoplasmic reticulum (ER) stress that is imposed by the secretory demands associated with environmental forces. In this role, the UPR has increasingly been shown to have crucial functions in immunity and inflammation. In this Review, we discuss the importance of the UPR in the development, differentiation, function and survival of immune cells in meeting the needs of an immune response. In addition, we review current insights into how the UPR is involved in complex chronic inflammatory diseases and, through its role in immune regulation, antitumour responses.
Figures
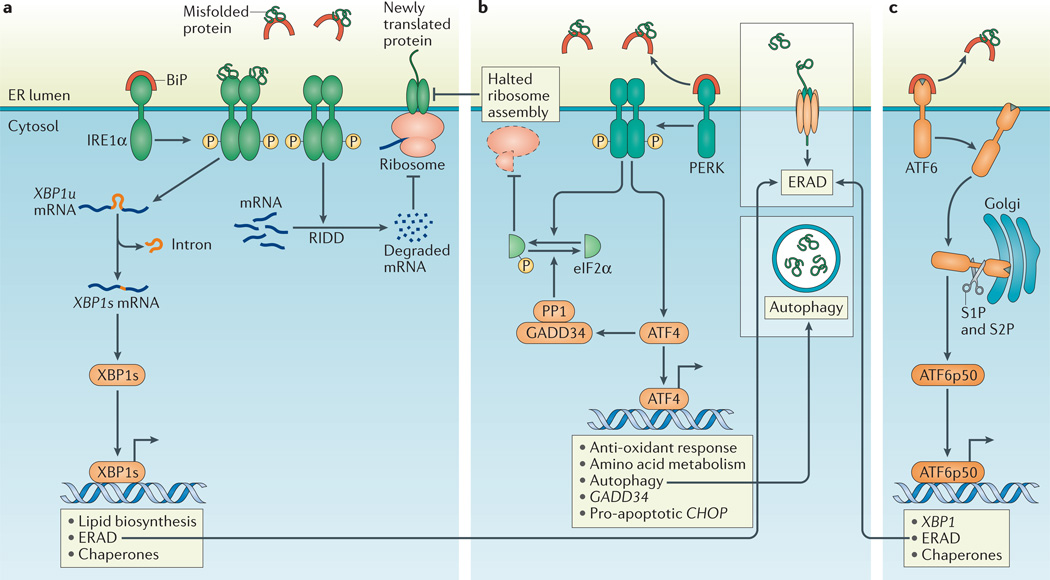
a | Dissociation of binding immunoglobulin protein (BiP) from inositol-requiring enzyme 1α (IRNEa1tuα). or dirct binding of misfolded proteins to IRE1α, activates the endoribonuclease domain of IRE1α, which non-conventionally splices an intron from unspliced X-box binding protein 1 (XBP1u) mRNA to produce XBP1s mRNA that encodes a potent transcriptional activator, XBP1s. Among the target genes of XBP1s are genes encoding proteins that increase the protein-folding capacity of the endoplasmic reticulum (ER) and that assist in the degradation of misfolded proteins by ER-associated degradation (ERAD; see inset box). In addition, the entry of newly synthesized proteins into the ER is limited by the degradation of mRNA through regulated IRE1α-dependent decay of mRNA (RIDD). b | PKR-like ER kinase (PERK)-dependent phosphorylation of eukaryotic translation initiation factor 2α (eIF2α) inhibits ribosome assembly, which causes a translational block and allows the cell to manage temporary ER stress. Activating transcription factor 4 (ATF4) escapes translation inhibition under ER stress conditions and induces the transcription of genes that promote survival, including those involved in compensatory autophagy (see inset box). Once ER stress is resolved, eIF2α is dephosphorylated by the GADD34–protein phosphatase 1 (PP1) complex to restore protein translation. However, if ER stress-induced damage is irreversible, the terminal unfolded protein response (UPR) is activated to induce apoptosis, mainly through C/EBP homologous protein (CHOP). c | Upon BiP dissociation from ATF6α during ER stress, ATF6α travels to the Golgi compartment where it is processed by the Golgi enzymes site 1 protease (S1P) and S2P to produce a cytosolic p50 fragment. ATF6p50 functions as a transcription factor that activates transcriptional programmes that increase ER capacity and protein folding, and that remove misfolded proteins from the ER for degradation (ERAD; see inset box).
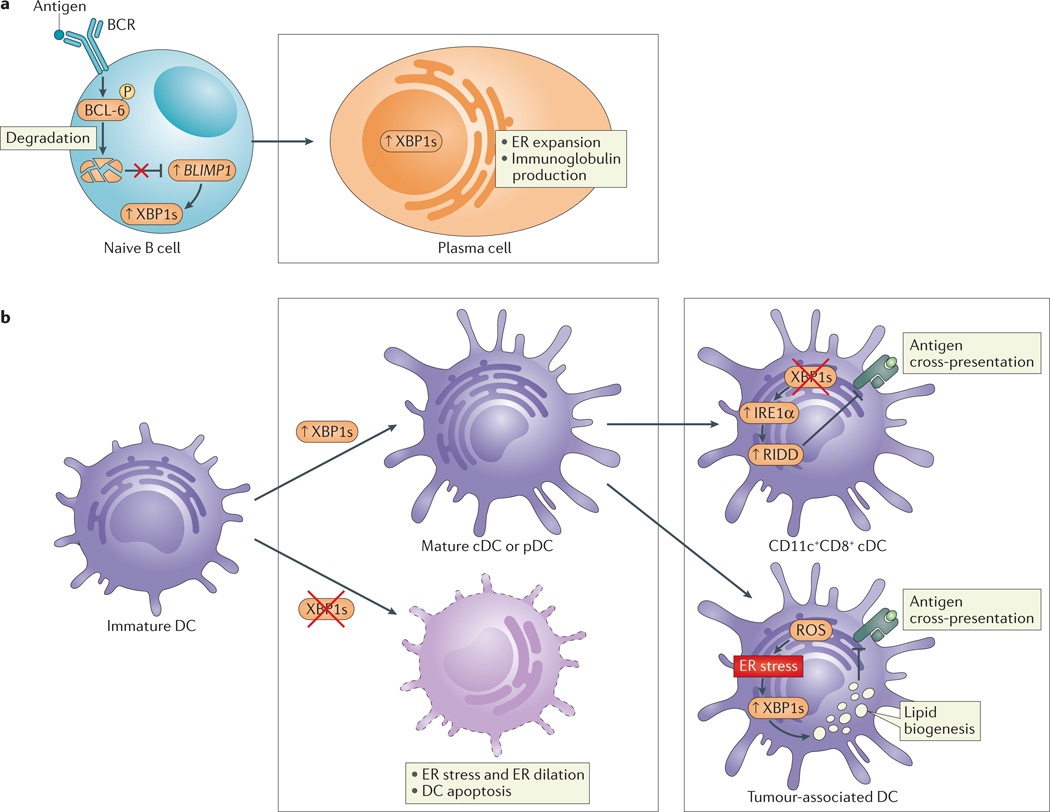
a | B cell receptor (BCR) ligation induces the phosphorylation of B cell lymphoma 6 (BCL-6) and its subsequent ubiquitylation and degradation. BCL-6 degradation derepresses B lymphocyte-induced maturation protein 1 (BLIMP1) in naive B cells to activate cellular programmes that are crucial for the development of plasma cells. These include activation of the unfolded protein response (UPR) and X-box binding protein 1 (XBP1) splicing, which is required for the expansion of the endoplasmic reticulum (ER) and increased protein (immunoglobulin) synthesis involved in plasma cell differentiation. b | XBP1 is crucial for the differentiation of conventional dendritic cells (cDCs) and plasmacytoid DCs (pDCs) from immature progenitors (left). Loss of XBP1 in progenitor cells abrogates maturation and decreases DC survival. XBP1 deletion in mature CD11c+ DCs results in inositol-requiring enzyme 1α (IRE1α) hyperactivation, leading to regulated IRE1α-dependent decay (RIDD)-dependent degradation of components of the MHC class I–mediated antigen cross-presentation machinery; as such, ER stress in DCs interferes with their function (right, top). By contrast, increased XBP1s in response to the production of reactive oxygen species (ROS) in tumour-associated DCs leads to augmented lipid biogenesis that is associated with the disruption of MHC class I–mediated antigen cross-presentation (right, bottom).
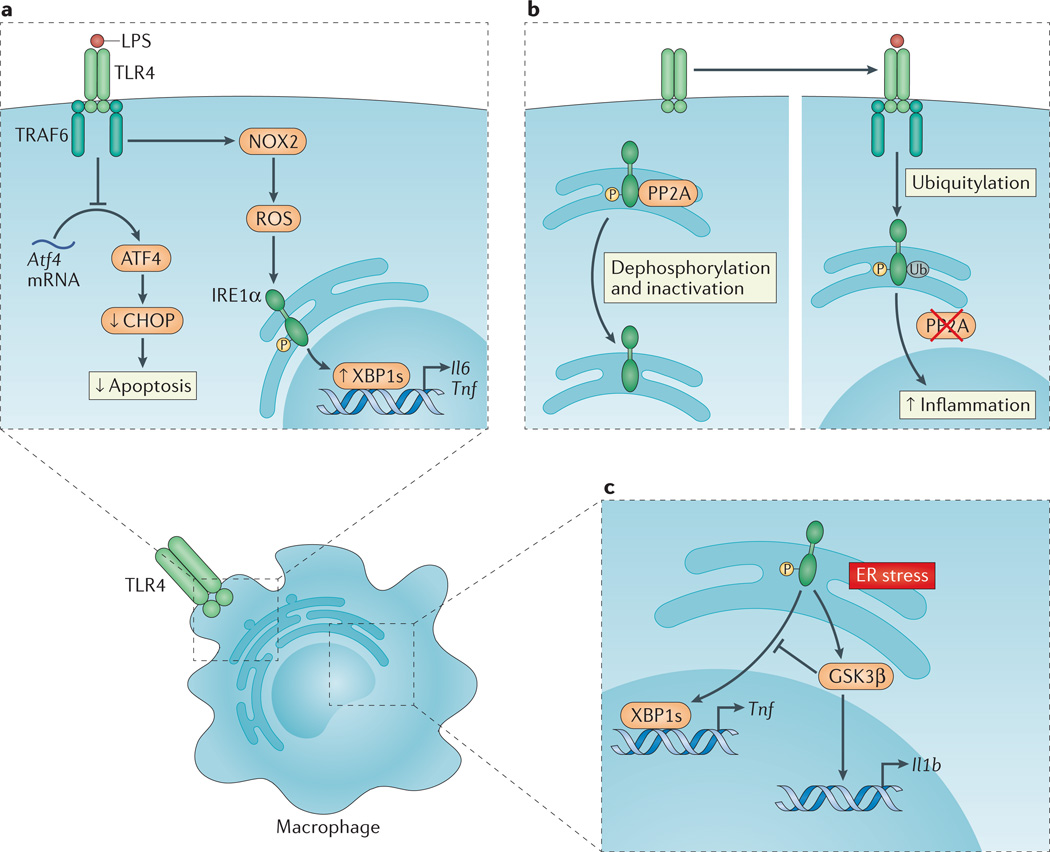
a | Upon Toll-like receptor (TLR) ligation, the inositol-requiring enzyme 1α (IRE1α) arm of the unfolded protein response (UPR) is activated through a mechanism that requires TNF receptor-associated factor 6 (TRAF6) recruitment to the TLR and reactive oxygen species (ROS) production by the NADPH oxidase NOX2. Spliced X-box binding protein 1 (XBP1s) functions as a transcription factor for the pro-inflammatory cytokines interleukin-6 (Il6) and tumour necrosis factor (Tnf). In a separate pathway, translation of activating transcription factor 4 (Atf4) mRNA is inhibited by TLR4 ligation, which decreases C/EBP homologous protein (CHOP) levels and apoptosis in activated macrophages. This favours macrophage survival and as such facilitates the immune response. b | In the absence of TLR signalling, phosphorylated IRE1α is subject to protein phosphatase 2A (PP2A)-mediated dephosphorylation and inactivation (left panel). Upon TLR ligation, however, TRAF6 interacts with and catalyses the ubiquitylation of IRE1α, which prevents PP2A–mediated dephosphorylation and inactivation of IRE1α, thereby amplifying inflammation (right panel). c | Endoplasmic reticulum (ER) stress-induced activation of IRE1α can induce Il1b transcription through glycogen synthase kinase 3β (GSK3β), which simultaneously inhibits Xbp1 splicing and thereby the transcription of XBP1s target genes, including Tnf.
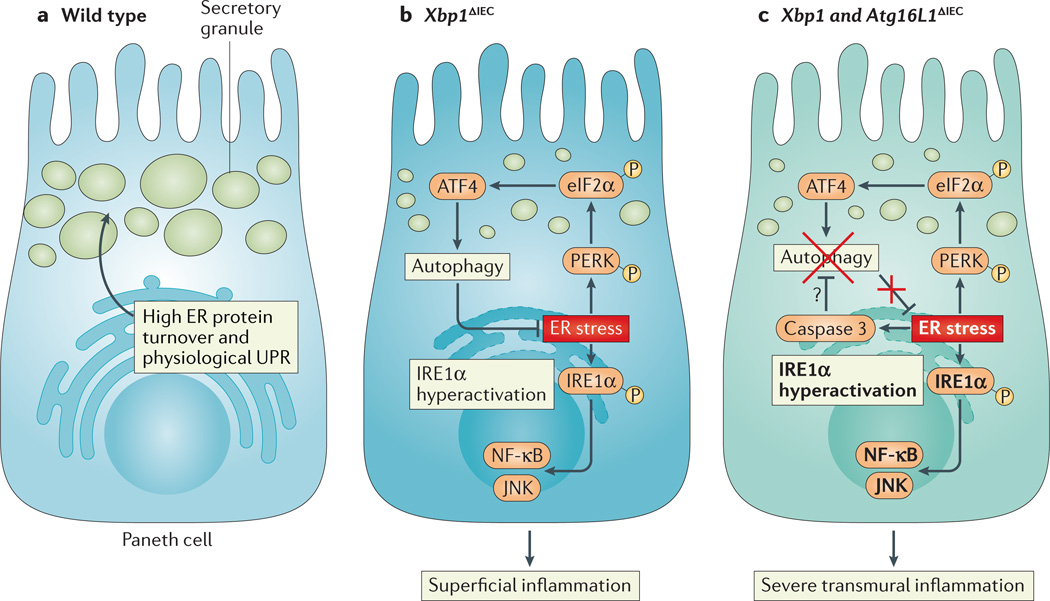
a | Intestinal epithelial cells, particularly Paneth cells, have a well-developed endoplasmic reticulum (ER) to manage their high secretory demands, including the production of antimicrobial proteins such as lysozyme and defensins. b | Deletion of X-box binding protein 1 (XBP1) in the small intestinal epithelium (Xbp1ΔIEC mice) induces ER stress, resulting in hypomorphic Paneth cells that have signs of inositol-requiring enzyme 1α (IRE1α) hyperactivation and downstream activation of nuclear factor-κB (NF-κB) and JUN N-terminal kinase (JNK). The induction of autophagy through the PKR-like ER kinase (PERK)-eukaryotic translation initiation factor 2α (eIF2α)-activating transcription factor 4 (ATF4) axis of the unfolded protein response (UPR) alleviates ER stress. c | Simultaneous deletion of XBP1 and autophagy-related 16 like 1 (ATG16L1) in the small intestinal epithelium thus further increases ER stress and ER stress-induced inflammation. Hypothetically, ER stress, which induces caspase 3, could be an important pathway responsible for the degradation of ATG16L1 in patients carrying the Crohn’s disease risk allele ATG16L1T300A, which is prone to caspase 3-mediated cleavage.
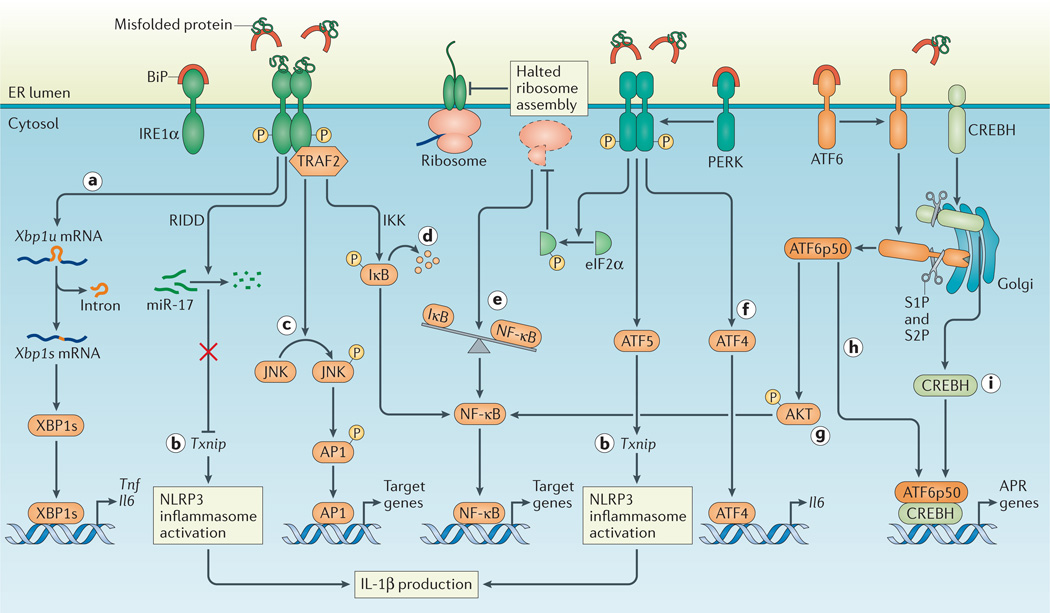
a | Inositol-requiring enzyme 1α (IRE1α) activation and subsequent splicing of X-box binding protein 1 (Xbp1) produces the transcription factor XBP1s that directly binds the promoters of tumour necrosis factor (Tnf) and interleukin-6 (Il6). b | Regulated IRE1α-dependent decay (RIDD)-dependent degradation of miR-17, which in unstressed conditions represses thioredoxin-interacting protein (Txnip), allows for increased TXNIP levels and NLRP3 inflammasome activation with upregulation of IL-1β. In addition, Txnip can be induced through the PKR-like ER kinase (PERK)-activating transcription factor 5 (ATF5) pathway to induce inflammasome activation. c | Activated IRE1α forms a complex with TNF receptor-associated factor 2 (TRAF2) to induce phosphorylation of Jun N-terminal kinase (JNK) and upregulation of pro-inflammatory genes through activated activator protein 1 (AP1). d | In addition, the IRE1α –TRAF2 complex recruits IκB kinase (IKK), and subsequent phosphorylation of IκB leads to its degradation, which releases nuclear factor-κB (NF-κB) for nuclear translocation. e | Translation attenuation by PERK-dependent phosphorylation of eukaryotic translation initiation factor 2α (eIF2α) results in decreased translation of both IκB and NF-κB. However, owing to the shorter half-life of IκB, the net result is an increased NF-κB to IκB ratio, which promotes inflammation. f | ATF4 directly binds the Il6 promoter. g | ATF6α induces NF-κB signalling via AKT phosphorylation. h,i | Site 1 protease (S1P)- and S2P–mediated cleavage of ATF6α and CREBH allows their cleavage fragments to translocate to the nucleus and induce genes of the acute phase response (APR). BiP, binding immunoglobulin protein.
References
Publication types
MeSH terms
Grants and funding
- R01 DK088199/DK/NIDDK NIH HHS/United States
- R01 DK051362/DK/NIDDK NIH HHS/United States
- P01 CA128814/CA/NCI NIH HHS/United States
- R01 DK053056/DK/NIDDK NIH HHS/United States
- P30 DK034854/DK/NIDDK NIH HHS/United States
- 260961/ERC_/European Research Council/International
- R01 DK042394/DK/NIDDK NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R56 DK053056/DK/NIDDK NIH HHS/United States
- R01 DK088227/DK/NIDDK NIH HHS/United States
- R37 DK044319/DK/NIDDK NIH HHS/United States
- 106260/Z/14/Z/WT_/Wellcome Trust/United Kingdom
- R37 DK042394/DK/NIDDK NIH HHS/United States
- R21 DK103183/DK/NIDDK NIH HHS/United States
- R01 DK044319/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
