

Segregated cholinergic transmission modulates dopamine neurons integrated in distinct functional circuits
- PMID: 27348215
- PMCID: PMC5086413
- DOI: 10.1038/nn.4335
Segregated cholinergic transmission modulates dopamine neurons integrated in distinct functional circuits
Abstract
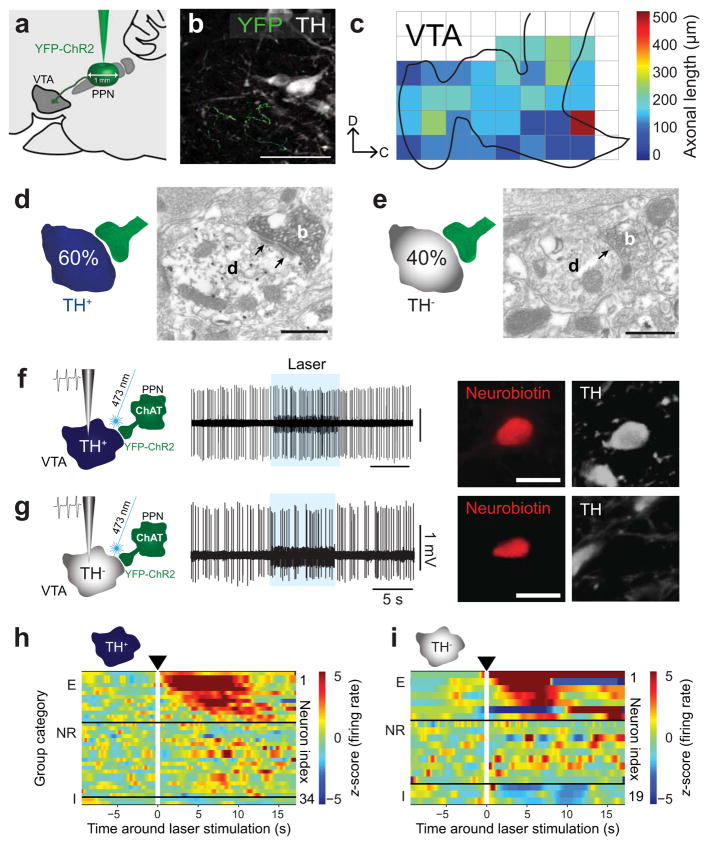
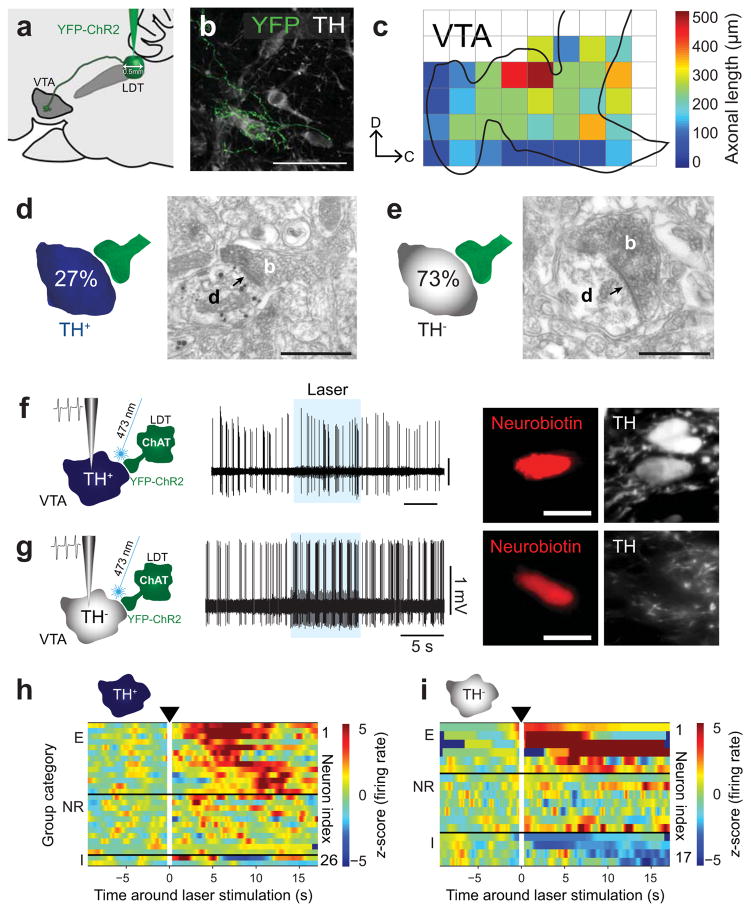
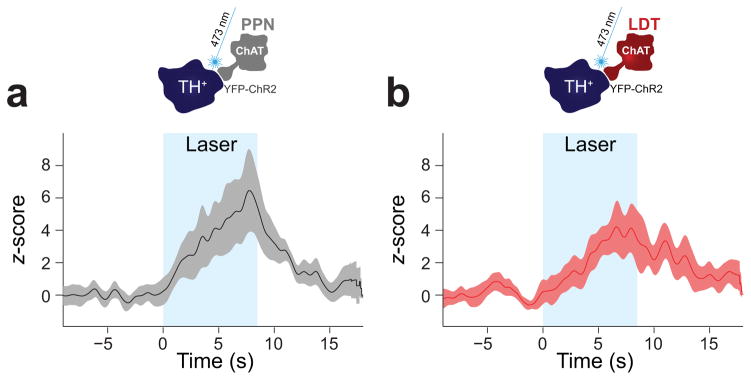
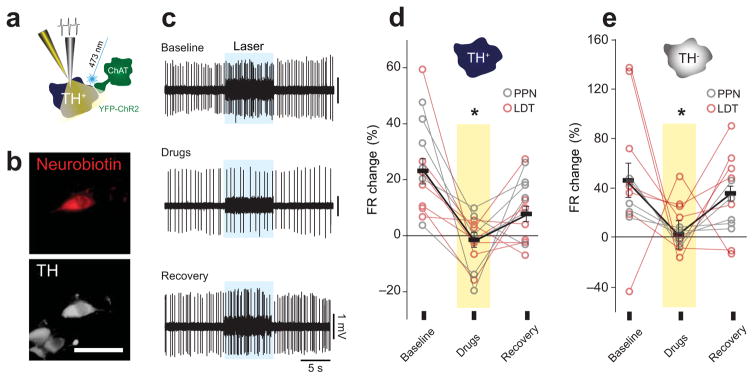
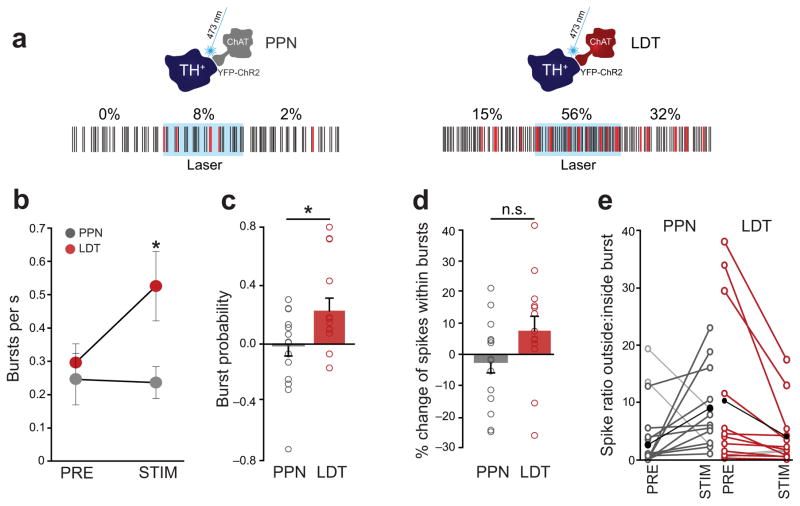
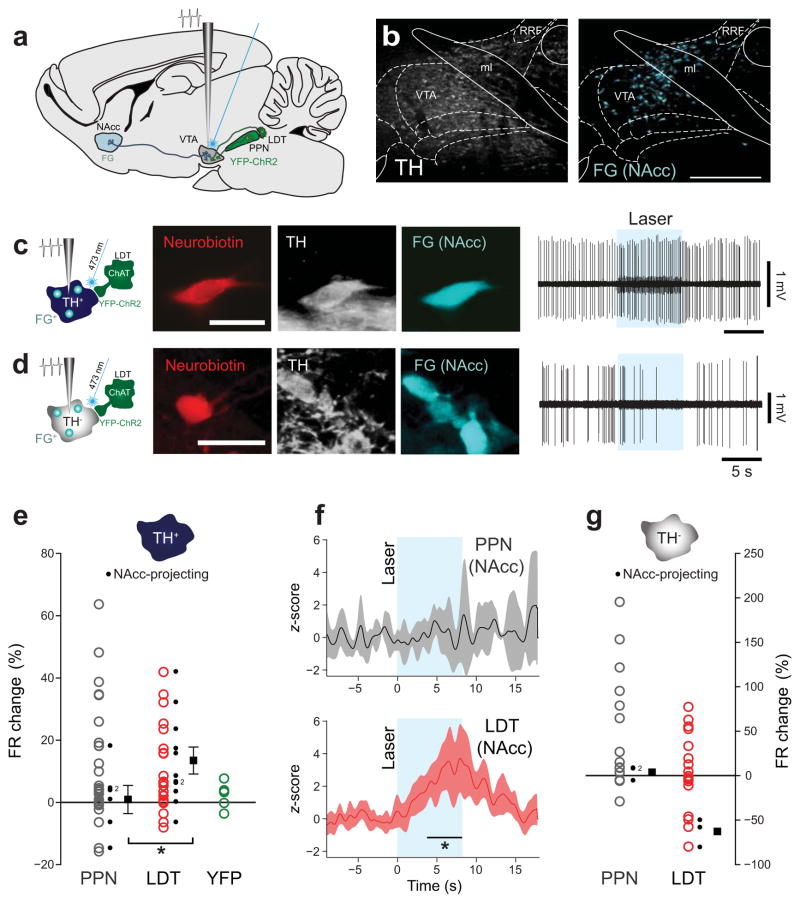
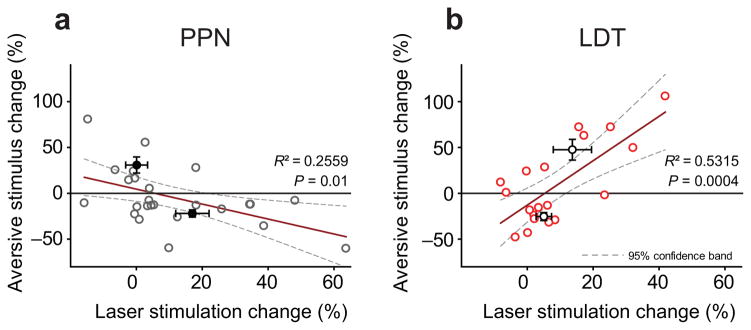
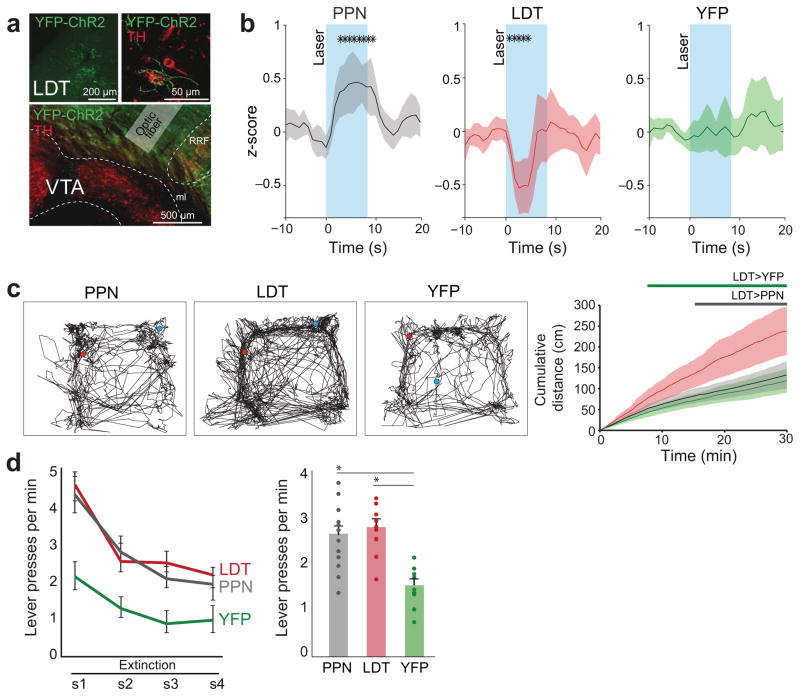
Dopamine neurons in the ventral tegmental area (VTA) receive cholinergic innervation from brainstem structures that are associated with either movement or reward. Whereas cholinergic neurons of the pedunculopontine nucleus (PPN) carry an associative/motor signal, those of the laterodorsal tegmental nucleus (LDT) convey limbic information. We used optogenetics and in vivo juxtacellular recording and labeling to examine the influence of brainstem cholinergic innervation of distinct neuronal subpopulations in the VTA. We found that LDT cholinergic axons selectively enhanced the bursting activity of mesolimbic dopamine neurons that were excited by aversive stimulation. In contrast, PPN cholinergic axons activated and changed the discharge properties of VTA neurons that were integrated in distinct functional circuits and were inhibited by aversive stimulation. Although both structures conveyed a reinforcing signal, they had opposite roles in locomotion. Our results demonstrate that two modes of cholinergic transmission operate in the VTA and segregate the neurons involved in different reward circuits.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Wise RA. Dopamine, learning and motivation. Nature Rev. 2004;5:483–494. - PubMed
-
- Montague PR, Hyman SE, Cohen JD. Computational roles for dopamine in behavioural control. Nature. 2004;431:760–767. - PubMed
-
- Joshua M, et al. Synchronization of midbrain dopaminergic neurons is enhanced by rewarding events. Neuron. 2009;62:695–704. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
