

Genomic analysis of local variation and recent evolution in Plasmodium vivax
- PMID: 27348299
- PMCID: PMC4966634
- DOI: 10.1038/ng.3599
Genomic analysis of local variation and recent evolution in Plasmodium vivax
Abstract
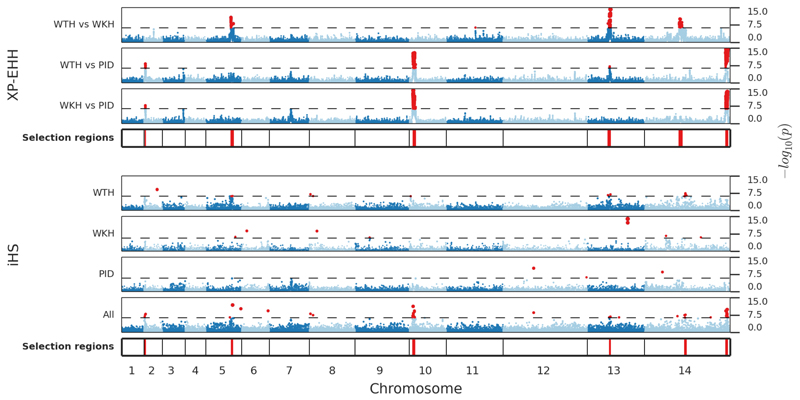
The widespread distribution and relapsing nature of Plasmodium vivax infection present major challenges for the elimination of malaria. To characterize the genetic diversity of this parasite in individual infections and across the population, we performed deep genome sequencing of >200 clinical samples collected across the Asia-Pacific region and analyzed data on >300,000 SNPs and nine regions of the genome with large copy number variations. Individual infections showed complex patterns of genetic structure, with variation not only in the number of dominant clones but also in their level of relatedness and inbreeding. At the population level, we observed strong signals of recent evolutionary selection both in known drug resistance genes and at new loci, and these varied markedly between geographical locations. These findings demonstrate a dynamic landscape of local evolutionary adaptation in the parasite population and provide a foundation for genomic surveillance to guide effective strategies for control and elimination of P. vivax.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
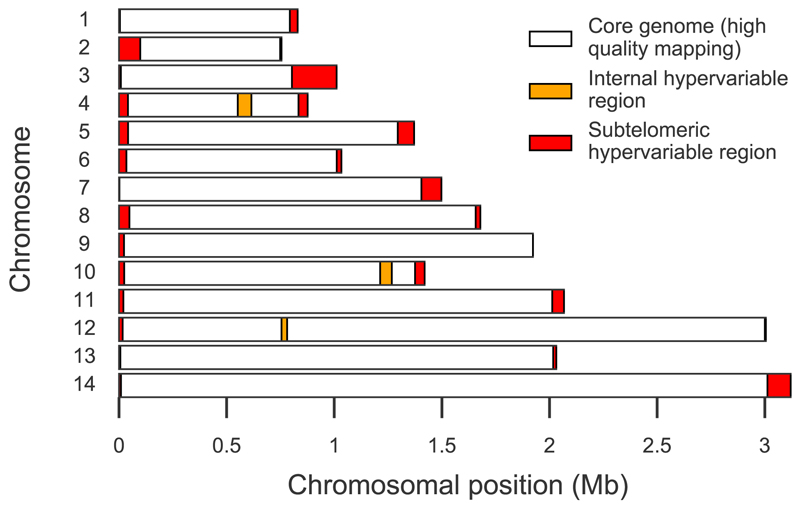
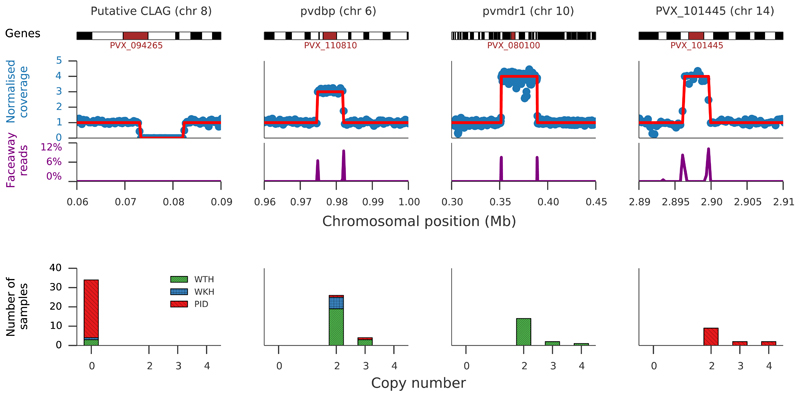
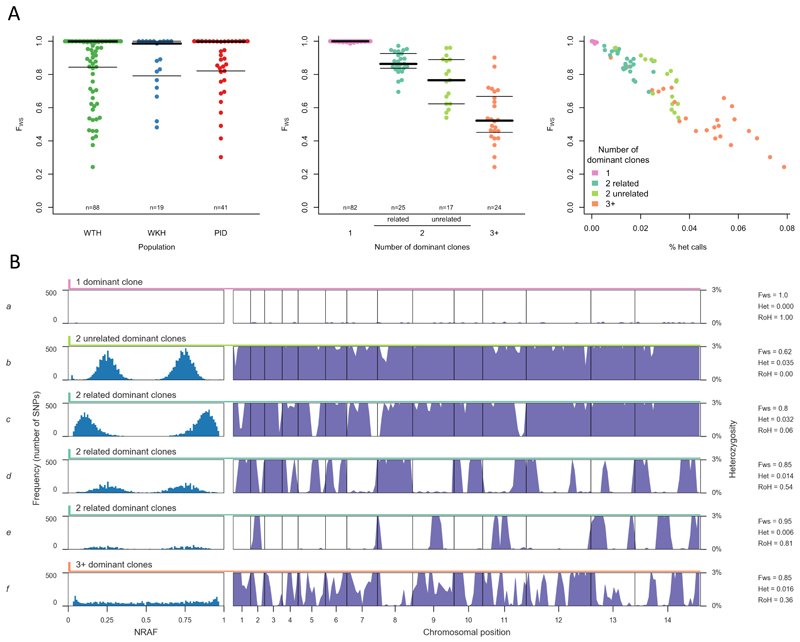
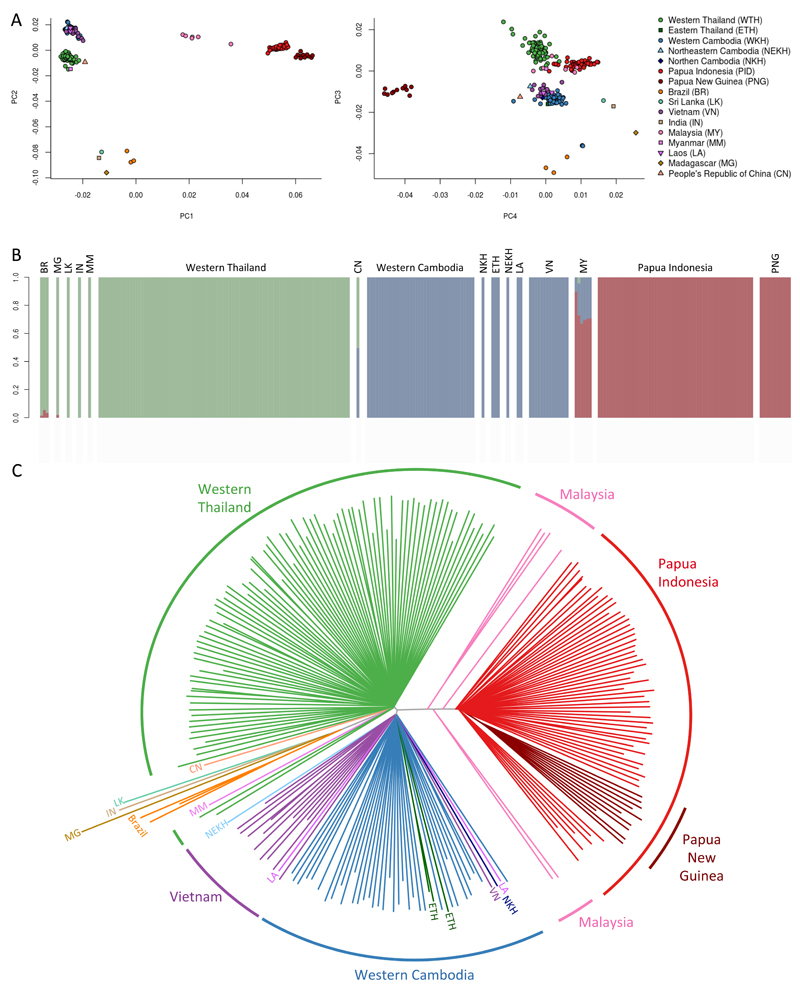
Comment in
-
An unsettling picture emerges from population genomic studies of Plasmodium vivax.Nat Genet. 2016 Jul 27;48(8):825-6. doi: 10.1038/ng.3630. Nat Genet. 2016. PMID: 27463397
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
