

Memory of cell shape biases stochastic fate decision-making despite mitotic rounding
- PMID: 27349214
- PMCID: PMC4931277
- DOI: 10.1038/ncomms11963
Memory of cell shape biases stochastic fate decision-making despite mitotic rounding
Abstract
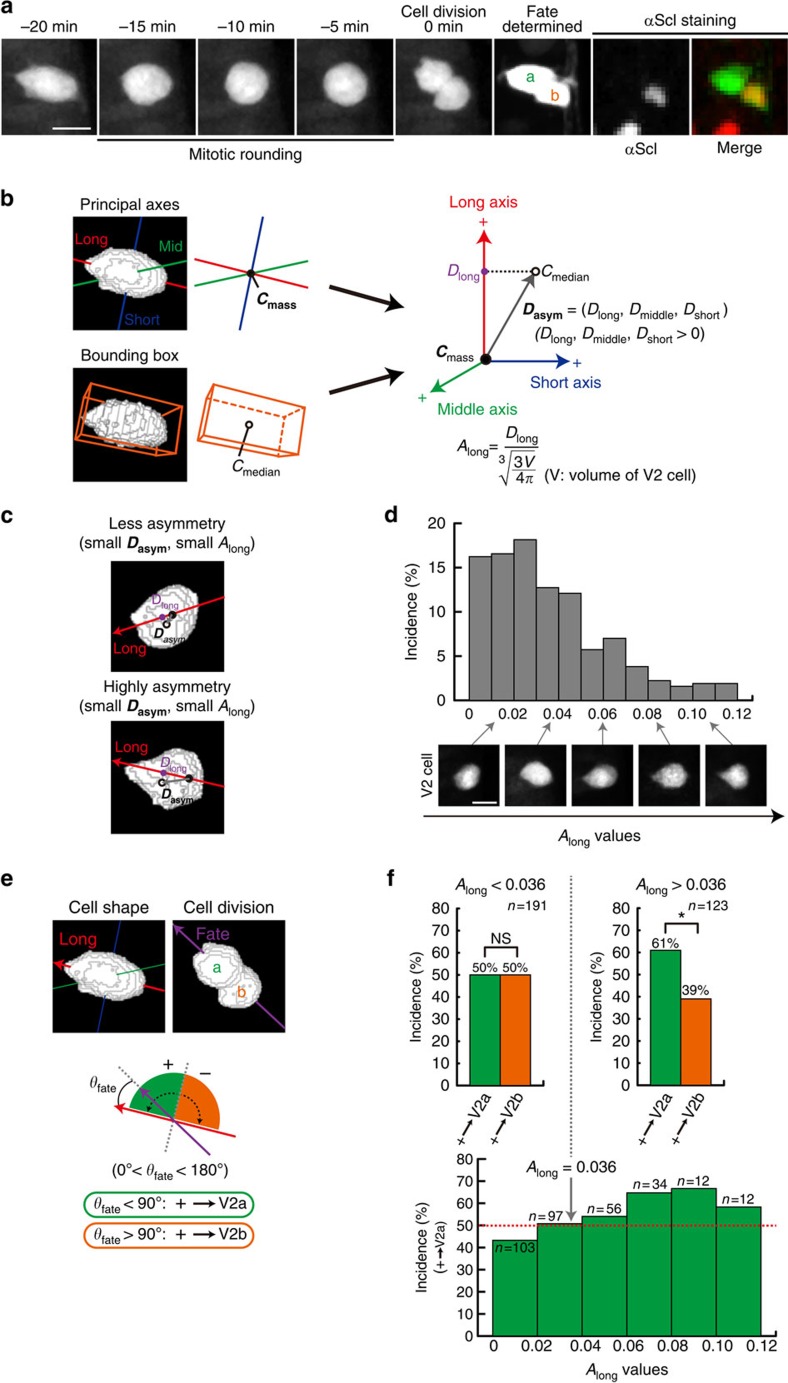
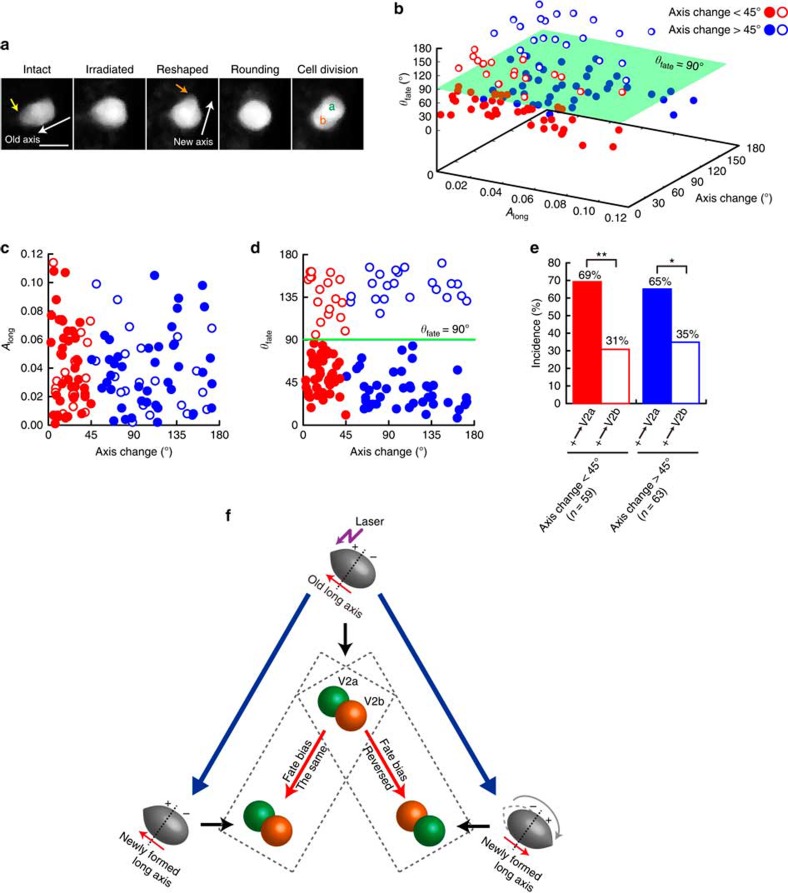
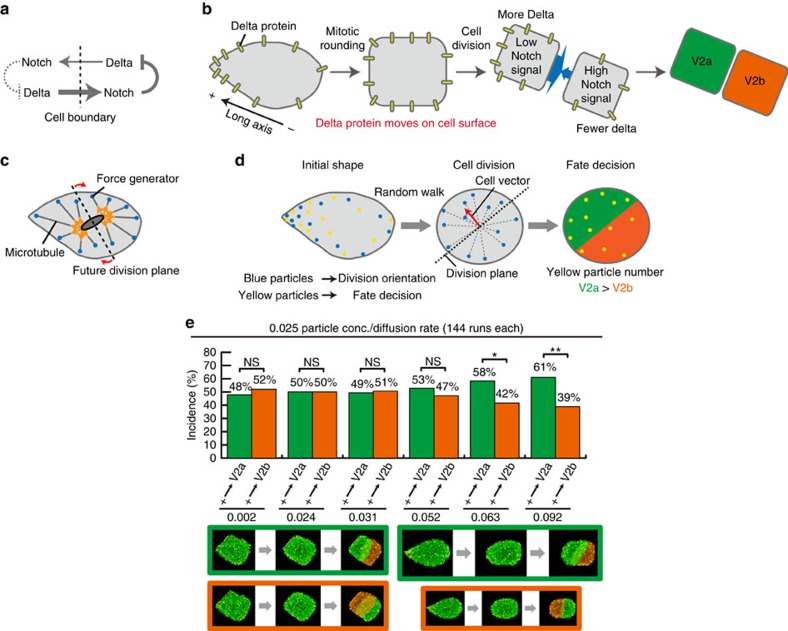
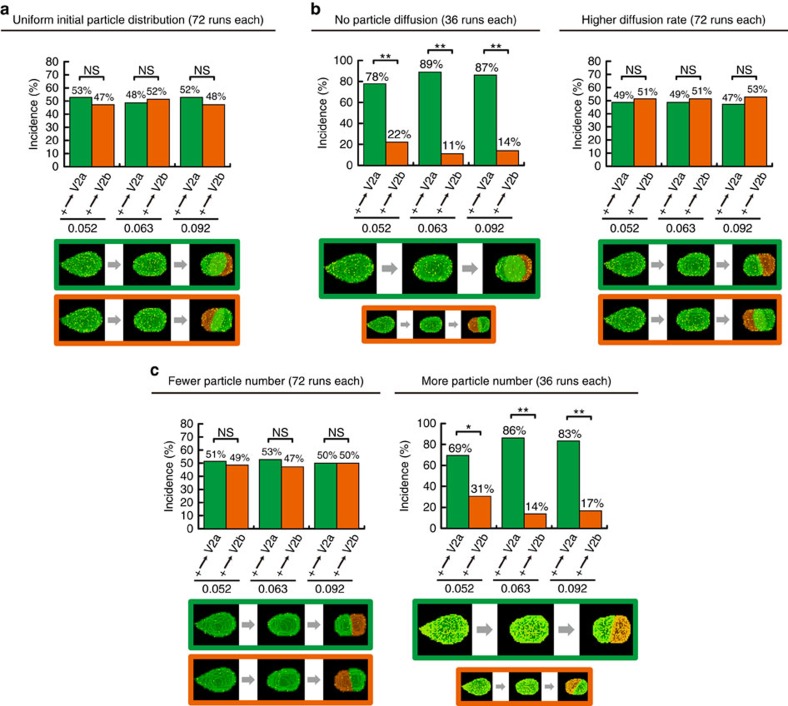
Cell shape influences function, and the current model suggests that such shape effect is transient. However, cells dynamically change their shapes, thus, the critical question is whether shape information remains influential on future cell function even after the original shape is lost. We address this question by integrating experimental and computational approaches. Quantitative live imaging of asymmetric cell-fate decision-making and their live shape manipulation demonstrates that cellular eccentricity of progenitor cell indeed biases stochastic fate decisions of daughter cells despite mitotic rounding. Modelling and simulation indicates that polarized localization of Delta protein instructs by the progenitor eccentricity is an origin of the bias. Simulation with varying parameters predicts that diffusion rate and abundance of Delta molecules quantitatively influence the bias. These predictions are experimentally validated by physical and genetic methods, showing that cells exploit a mechanism reported herein to influence their future fates based on their past shape despite dynamic shape changes.
Figures
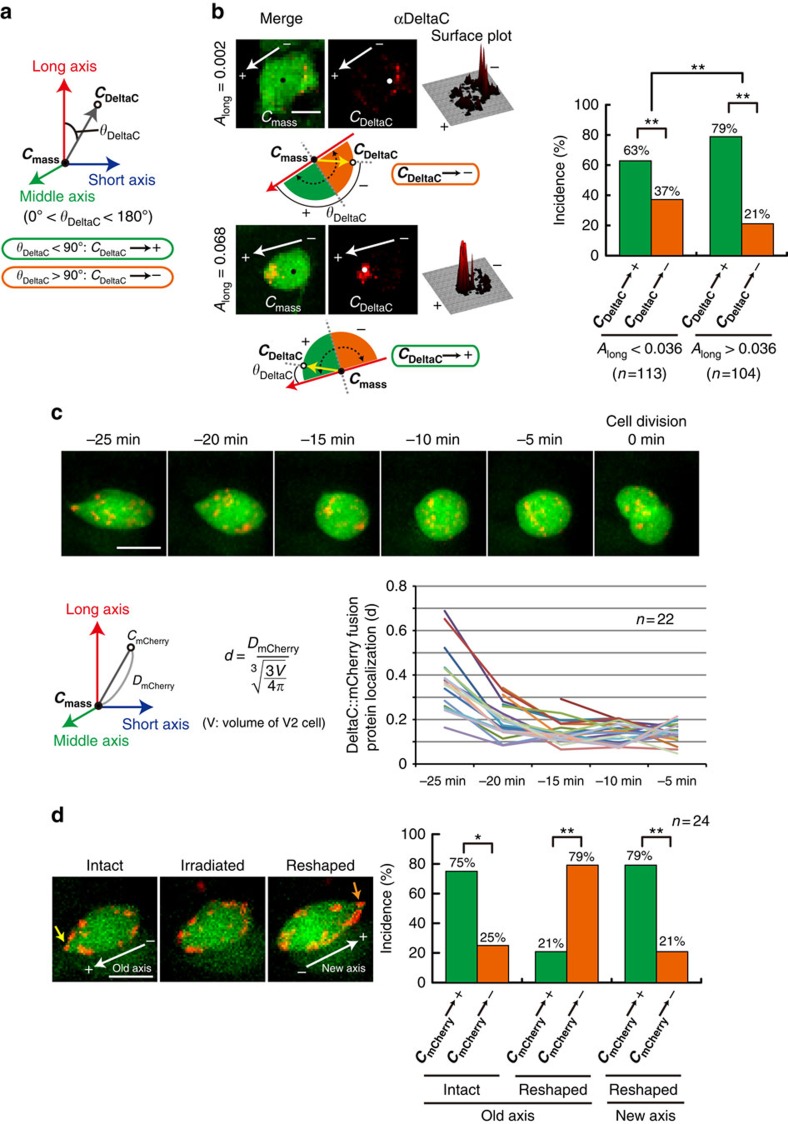
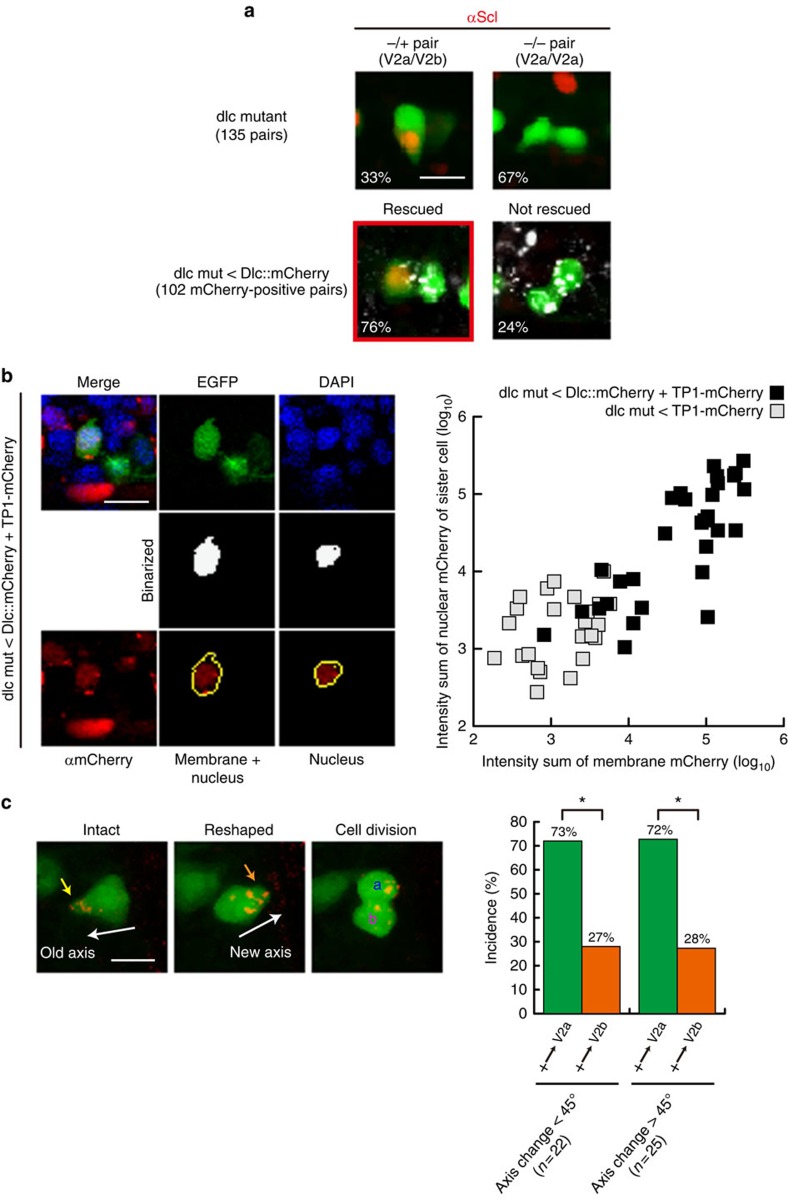
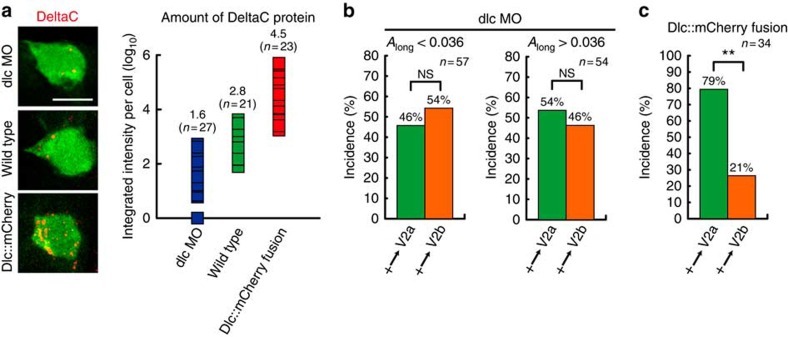
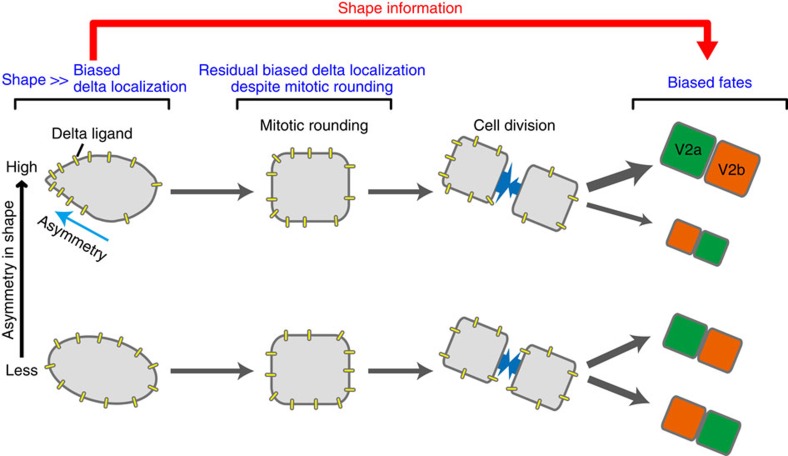
Comment in
-
Shaping the cell fate.Cell Cycle. 2017 Jan 17;16(2):149-150. doi: 10.1080/15384101.2016.1241603. Epub 2016 Oct 13. Cell Cycle. 2017. PMID: 27736341 Free PMC article. No abstract available.
References
-
- Hertwig O. Das Problem der Fefrunchtung une der isotropie des Eies, eine Theory der Vererbung. Jenaische Zeitschrift für Naturwissenschaft 18, 276–318 (1884).
-
- Minc N. & Piel M. Predicting division plane position and orientation. Trends Cell Biol. 22, 193–200 (2012). - PubMed
-
- Folkman J. & Moscona A. Role of cell shape in growth control. Nature 273, 345–349 (1978). - PubMed
-
- Meyers J., Craig J. & Odde D. J. Potential for control of signaling pathways via cell size and shape. Curr. Biol. 16, 1685–1693 (2006). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
