

Targeting Alpha Toxin and ClfA with a Multimechanistic Monoclonal-Antibody-Based Approach for Prophylaxis of Serious Staphylococcus aureus Disease
- PMID: 27353753
- PMCID: PMC4937210
- DOI: 10.1128/mBio.00528-16
Targeting Alpha Toxin and ClfA with a Multimechanistic Monoclonal-Antibody-Based Approach for Prophylaxis of Serious Staphylococcus aureus Disease
Abstract
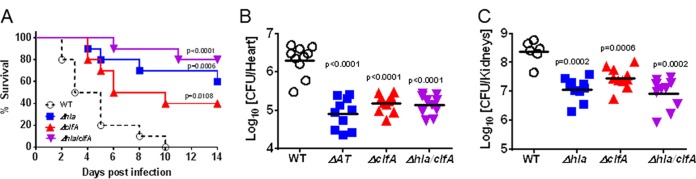
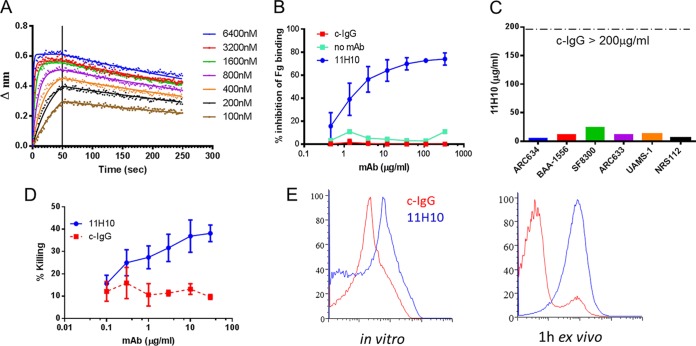
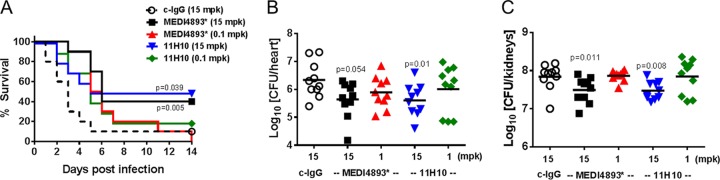
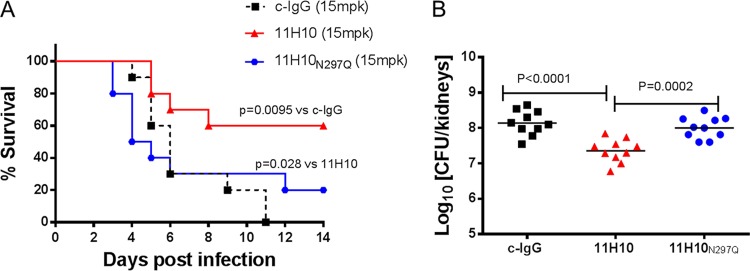
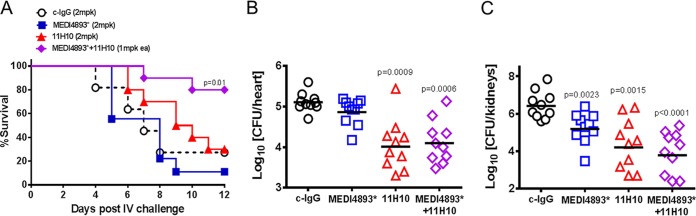
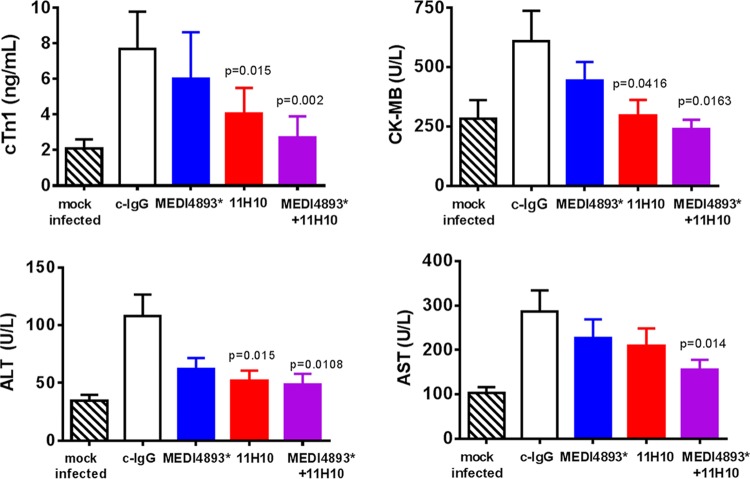
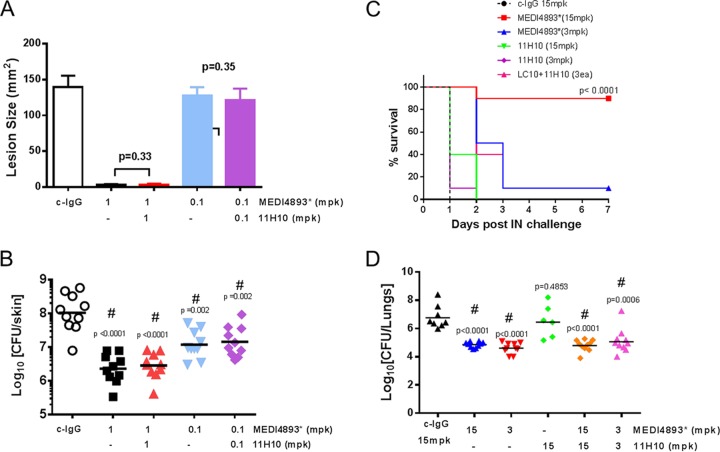
Staphylococcus aureus produces numerous virulence factors, each contributing different mechanisms to bacterial pathogenesis in a spectrum of diseases. Alpha toxin (AT), a cytolytic pore-forming toxin, plays a key role in skin and soft tissue infections and pneumonia, and a human anti-AT monoclonal antibody (MAb), MEDI4893*, has been shown to reduce disease severity in dermonecrosis and pneumonia infection models. However, interstrain diversity and the complex pathogenesis of S. aureus bloodstream infections suggests that MEDI4893* alone may not provide adequate protection against S. aureus sepsis. Clumping factor A (ClfA), a fibrinogen binding protein, is an important virulence factor facilitating S. aureus bloodstream infections. Herein, we report on the identification of a high-affinity anti-ClfA MAb, 11H10, that inhibits ClfA binding to fibrinogen, prevents bacterial agglutination in human plasma, and promotes opsonophagocytic bacterial killing (OPK). 11H10 prophylaxis reduced disease severity in a mouse bacteremia model and was dependent on Fc effector function and OPK. Additionally, prophylaxis with 11H10 in combination with MEDI4893* provided enhanced strain coverage in this model and increased survival compared to that obtained with the individual MAbs. The MAb combination also reduced disease severity in murine dermonecrosis and pneumonia models, with activity similar to that of MEDI4893* alone. These results indicate that an MAb combination targeting multiple virulence factors provides benefit over a single MAb neutralizing one virulence mechanism by providing improved efficacy, broader strain coverage, and protection against multiple infection pathologies.
Importance: Alternative strategies to broad-spectrum antibiotics are required to combat the antibiotic resistance epidemic. Previous attempts at active or passive immunization against Staphylococcus aureus targeting single antigens have failed in clinical trials despite positive preclinical data. To provide broad disease and isolate coverage, an effective immunization strategy likely must target multiple virulence mechanisms of the pathogen. Herein, we tested a multimechanistic MAb combination targeting alpha toxin (AT) and clumping factor A (ClfA) that neutralizes AT-mediated cytotoxicity, blocks fibrinogen binding by ClfA, prevents bacterial agglutination, targets the bacteria for opsonophagocytic killing, and provides broad isolate coverage in a lethal-bacteremia model. Although each MAb alone was effective in bacteremia against some individual isolates, the MAb combination provided improved protection against other isolates. These results illustrate the importance of targeting multiple virulence mechanisms and highlight the potential for an MAb combination targeting AT and ClfA to effectively prevent S. aureus disease.
Copyright © 2016 Tkaczyk et al.
Figures
References
-
- Laupland KB, Lyytikäinen O, Søgaard M, Kennedy KJ, Knudsen JD, Ostergaard C, Galbraith JC, Valiquette L, Jacobsson G, Collignon P, Schønheyder HC, International Bacteremia Surveillance Collaborative . 2013. The changing epidemiology of Staphylococcus aureus bloodstream infection: a multinational population-based surveillance study. Clin Microbiol Infect 19:465–471. doi: 10.1111/j.1469-0691.2012.03903.x. - DOI - PubMed
-
- Bagnoli F, Fontana MR, Soldaini E, Mishra RP, Fiaschi L, Cartocci E, Nardi-Dei V, Ruggiero P, Nosari S, De Falco MG, Lofano G, Marchi S, Galletti B, Mariotti P, Bacconi M, Torre A, Maccari S, Scarselli M, Rinaudo CD, Inoshima N, Savino S, Mori E, Rossi-Paccani S, Baudner B, Pallaoro M, Swennen E, Petracca R, Brettoni C, Liberatori S, Norais N, Monaci E, Bubeck Wardenburg J, Schneewind O, O’Hagan DT, Valiante NM, Bensi G, Bertholet S, De Gregorio E, Rappuoli R, Grandi G. 2015. Vaccine composition formulated with a novel TLR7-dependent adjuvant induces high and broad protection against Staphylococcus aureus. Proc Natl Acad Sci U S A 112:3680–3685. doi: 10.1073/pnas.1424924112. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
