

Chromosome-level assembly of Arabidopsis thaliana Ler reveals the extent of translocation and inversion polymorphisms
- PMID: 27354520
- PMCID: PMC4948326
- DOI: 10.1073/pnas.1607532113
Chromosome-level assembly of Arabidopsis thaliana Ler reveals the extent of translocation and inversion polymorphisms
Abstract
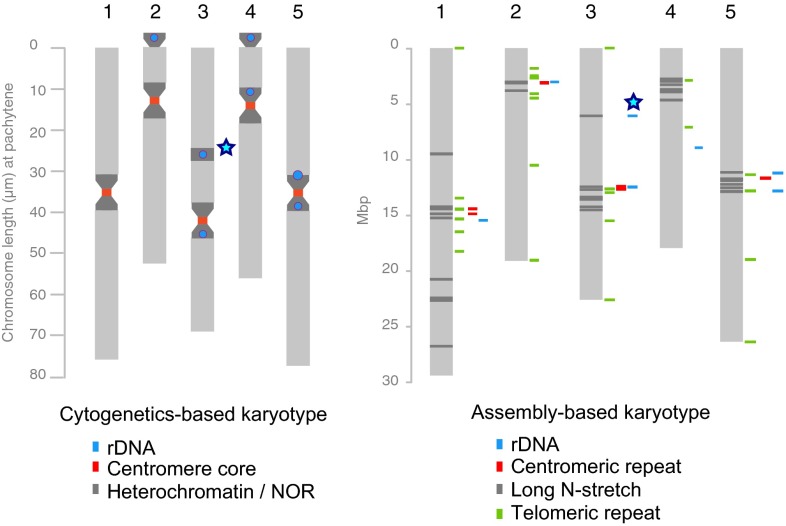
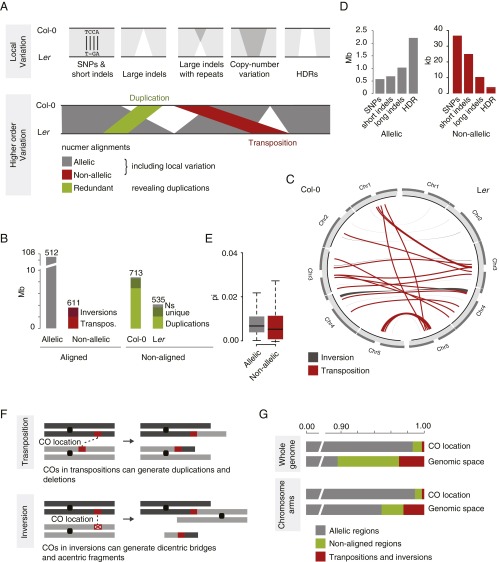
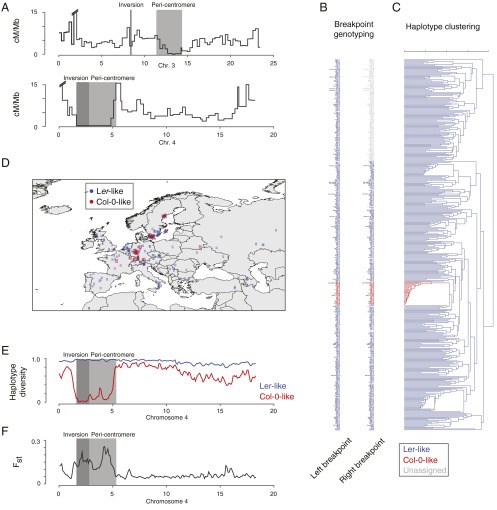
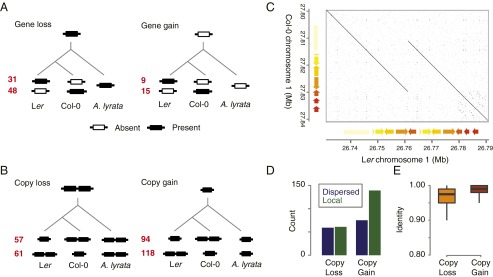
Resequencing or reference-based assemblies reveal large parts of the small-scale sequence variation. However, they typically fail to separate such local variation into colinear and rearranged variation, because they usually do not recover the complement of large-scale rearrangements, including transpositions and inversions. Besides the availability of hundreds of genomes of diverse Arabidopsis thaliana accessions, there is so far only one full-length assembled genome: the reference sequence. We have assembled 117 Mb of the A. thaliana Landsberg erecta (Ler) genome into five chromosome-equivalent sequences using a combination of short Illumina reads, long PacBio reads, and linkage information. Whole-genome comparison against the reference sequence revealed 564 transpositions and 47 inversions comprising ∼3.6 Mb, in addition to 4.1 Mb of nonreference sequence, mostly originating from duplications. Although rearranged regions are not different in local divergence from colinear regions, they are drastically depleted for meiotic recombination in heterozygotes. Using a 1.2-Mb inversion as an example, we show that such rearrangement-mediated reduction of meiotic recombination can lead to genetically isolated haplotypes in the worldwide population of A. thaliana Moreover, we found 105 single-copy genes, which were only present in the reference sequence or the Ler assembly, and 334 single-copy orthologs, which showed an additional copy in only one of the genomes. To our knowledge, this work gives first insights into the degree and type of variation, which will be revealed once complete assemblies will replace resequencing or other reference-dependent methods.
Keywords: Arabidopsis; PacBio sequencing; de novo assembly; gene absence/presence polymorphisms; inversions.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Rédei GP. Single loci heterosis. Z Vererbungsl. 1962;93(1):164–170.
-
- Rédei GP. A heuristic glance at the past of Arabidopsis genetics. In: Koncz C, Chua NH, Schell J, editors. Methods in Arabidopsis Research. World Scientific; Singapore: 1992. pp. 1–15.
-
- Fransz P, et al. Cytogenetics for the model system Arabidopsis thaliana. Plant J. 1998;13(6):867–876. - PubMed
-
- Fransz PF, et al. Integrated cytogenetic map of chromosome arm 4S of A. thaliana: Structural organization of heterochromatic knob and centromere region. Cell. 2000;100(3):367–376. - PubMed
Publication types
MeSH terms
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
