

Bacterial lipids activate, synergize, and inhibit a developmental switch in choanoflagellates
- PMID: 27354530
- PMCID: PMC4948368
- DOI: 10.1073/pnas.1605015113
Bacterial lipids activate, synergize, and inhibit a developmental switch in choanoflagellates
Abstract
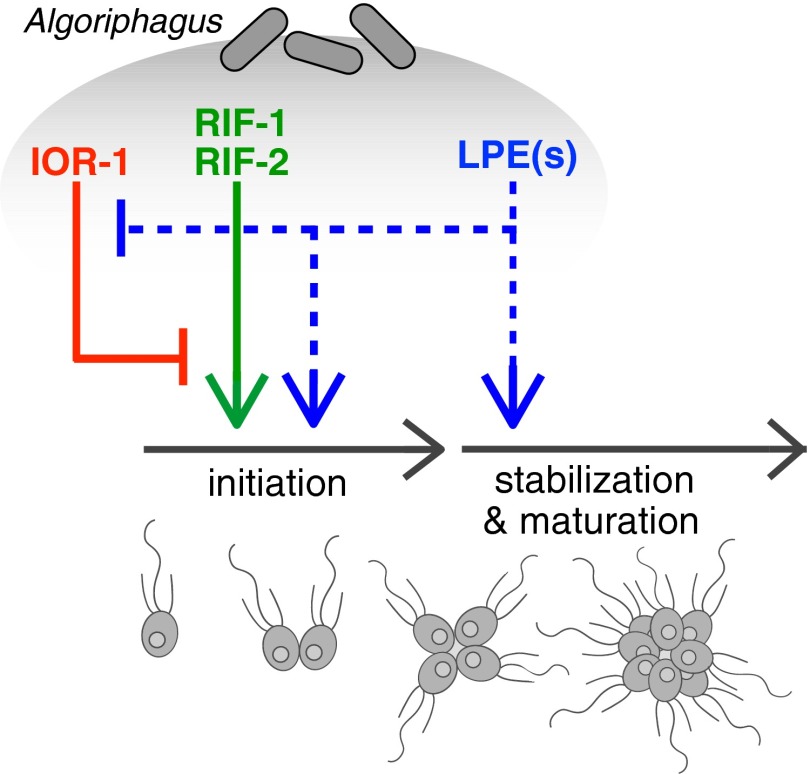
In choanoflagellates, the closest living relatives of animals, multicellular rosette development is regulated by environmental bacteria. The simplicity of this evolutionarily relevant interaction provides an opportunity to identify the molecules and regulatory logic underpinning bacterial regulation of development. We find that the rosette-inducing bacterium Algoriphagus machipongonensis produces three structurally divergent classes of bioactive lipids that, together, activate, enhance, and inhibit rosette development in the choanoflagellate Salpingoeca rosetta. One class of molecules, the lysophosphatidylethanolamines (LPEs), elicits no response on its own but synergizes with activating sulfonolipid rosette-inducing factors (RIFs) to recapitulate the full bioactivity of live Algoriphagus. LPEs, although ubiquitous in bacteria and eukaryotes, have not previously been implicated in the regulation of a host-microbe interaction. This study reveals that multiple bacterially produced lipids converge to activate, enhance, and inhibit multicellular development in a choanoflagellate.
Keywords: bacteria; choanoflagellates; host–microbe; multicellularity; sulfonolipid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Carroll SB. Chance and necessity: The evolution of morphological complexity and diversity. Nature. 2001;409(6823):1102–1109. - PubMed
-
- Tyler S. Epithelium: The primary building block for metazoan complexity. Integr Comp Biol. 2003;43(1):55–63. - PubMed
-
- King N. The unicellular ancestry of animal development. Dev Cell. 2004;7(3):313–325. - PubMed
-
- McFall-Ngai M. The development of cooperative associations between animals and bacteria: Establishing détente among domains. Am Zool. 1998;38(4):592–608.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
