

Systemic desensitization through TRPA1 channels by capsazepine and mustard oil - a novel strategy against inflammation and pain
- PMID: 27356469
- PMCID: PMC4928060
- DOI: 10.1038/srep28621
Systemic desensitization through TRPA1 channels by capsazepine and mustard oil - a novel strategy against inflammation and pain
Abstract
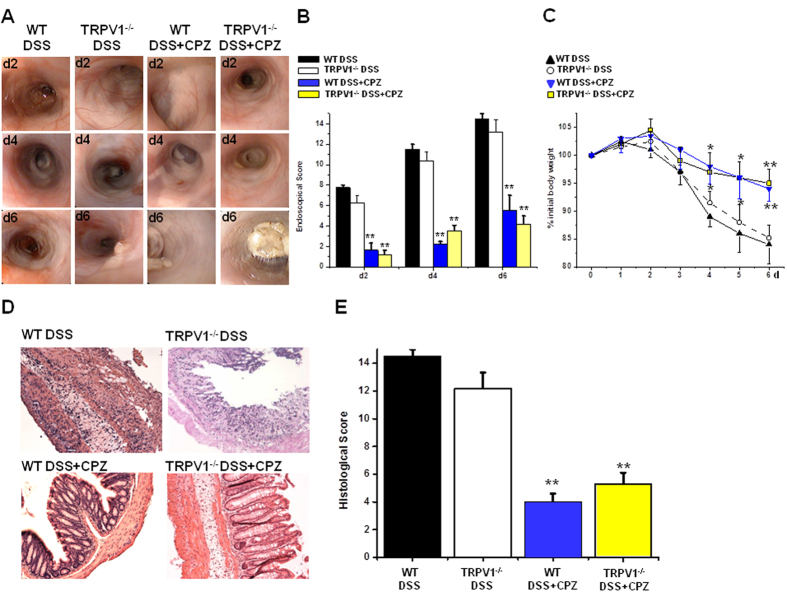
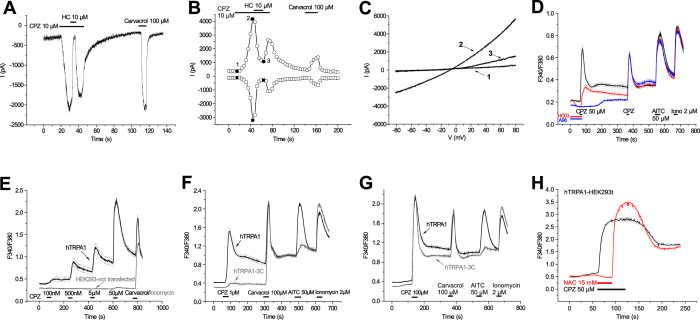
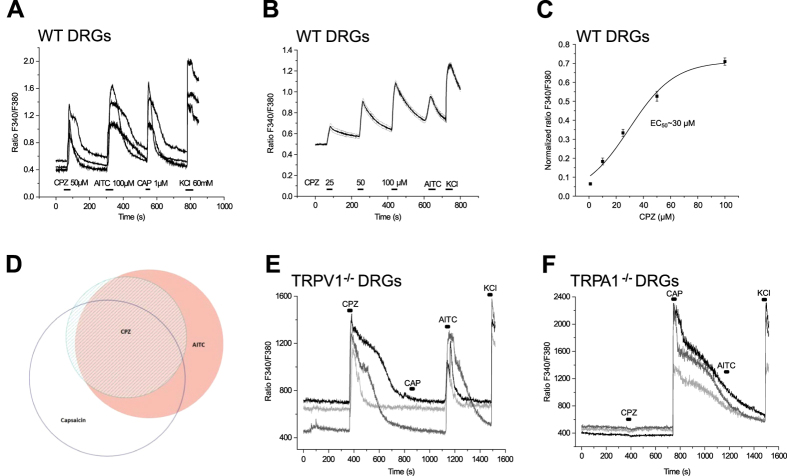
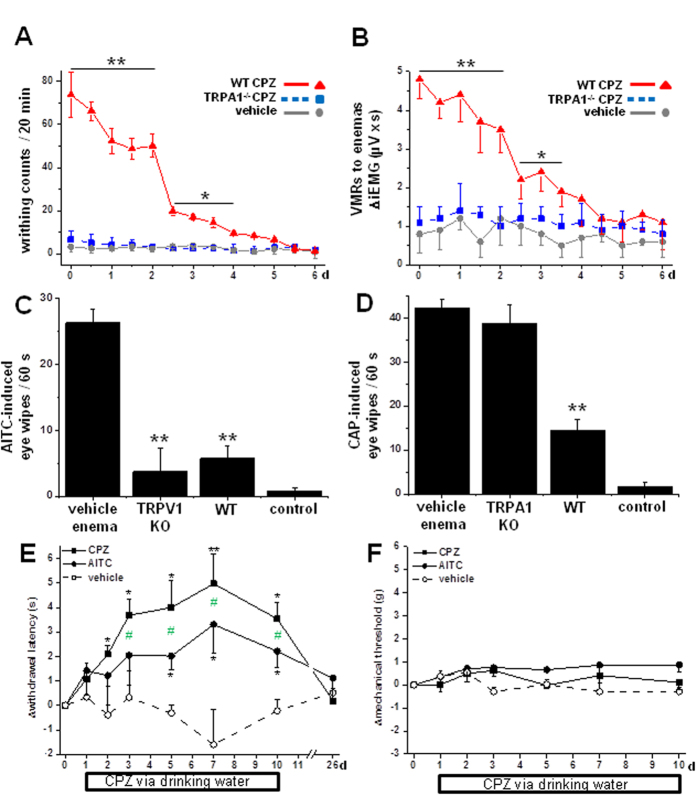
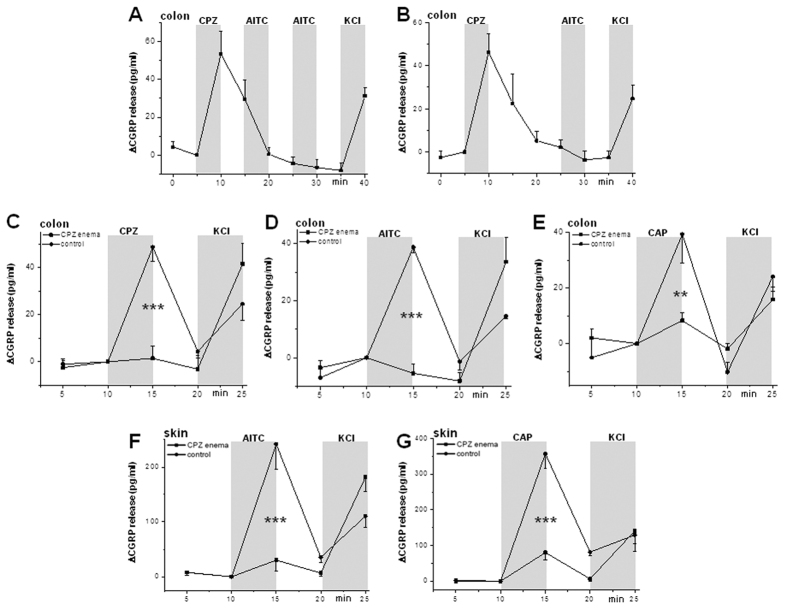
We demonstrate a novel dual strategy against inflammation and pain through body-wide desensitization of nociceptors via TRPA1. Attenuation of experimental colitis by capsazepine (CPZ) has long been attributed to its antagonistic action on TRPV1 and associated inhibition of neurogenic inflammation. In contrast, we found that CPZ exerts its anti-inflammatory effects via profound desensitization of TRPA1. Micromolar CPZ induced calcium influx in isolated dorsal root ganglion (DRG) neurons from wild-type (WT) but not TRPA1-deficient mice. CPZ-induced calcium transients in human TRPA1-expressing HEK293t cells were blocked by the selective TRPA1 antagonists HC 030031 and A967079 and involved three cysteine residues in the N-terminal domain. Intriguingly, both colonic enemas and drinking water with CPZ led to profound systemic hypoalgesia in WT and TRPV1(-/-) but not TRPA1(-/-) mice. These findings may guide the development of a novel class of disease-modifying drugs with anti-inflammatory and anti-nociceptive effects.
Figures
References
-
- Julius D. TRP channels and pain. Annu. Rev. Cell. Dev. Biol. 29, 355–384 (2013). - PubMed
-
- Caterina M. J. et al. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 288, 306–313 (2000). - PubMed
-
- Davis J. B. et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 405, 183–187 (2000). - PubMed
-
- Macpherson L. J. et al. Noxious compounds activate TRPA1 ion channels through covalent modification of cysteines. Nature 445, 541–545 (2007). - PubMed
-
- Petho G. & Reeh P. W. Sensory and signaling mechanisms of bradykinin, eicosanoids, platelet-activating factor, and nitric oxide in peripheral nociceptors. Physiol. Rev. 92, 1699–1775 (2012). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
