

Expansion of amphibian intronless interferons revises the paradigm for interferon evolution and functional diversity
- PMID: 27356970
- PMCID: PMC4928184
- DOI: 10.1038/srep29072
Expansion of amphibian intronless interferons revises the paradigm for interferon evolution and functional diversity
Abstract
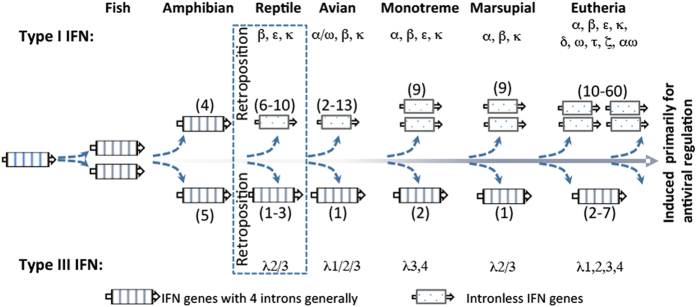
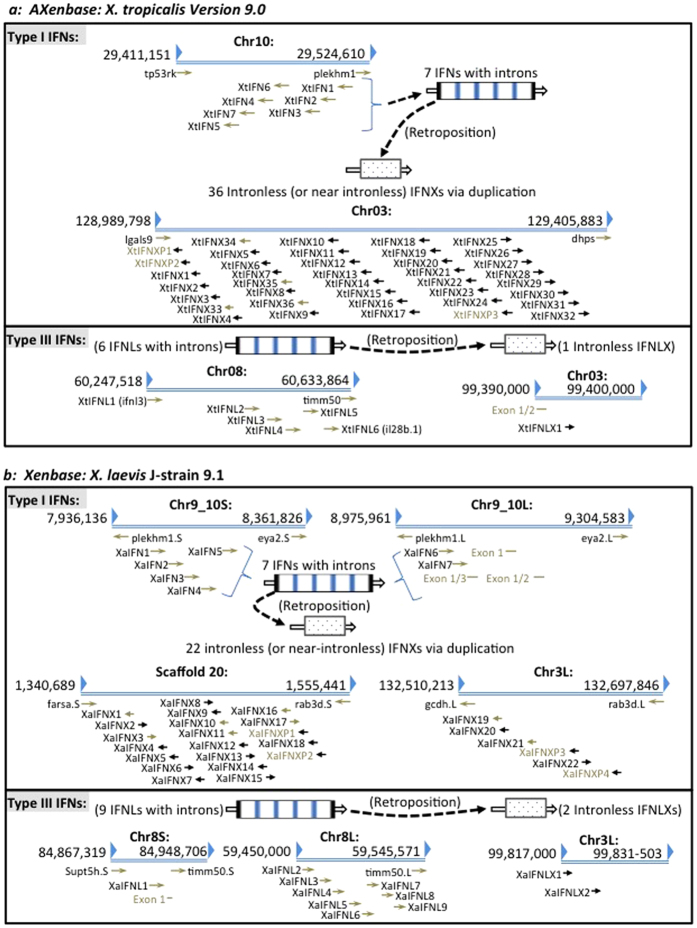
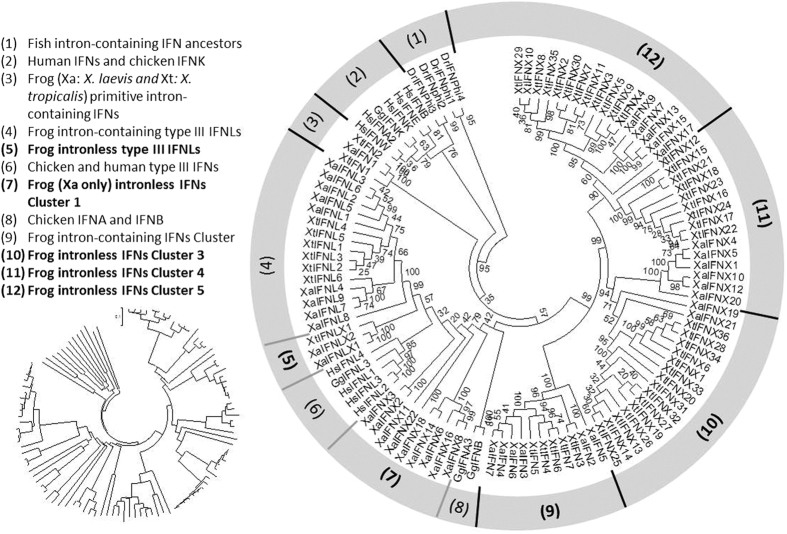
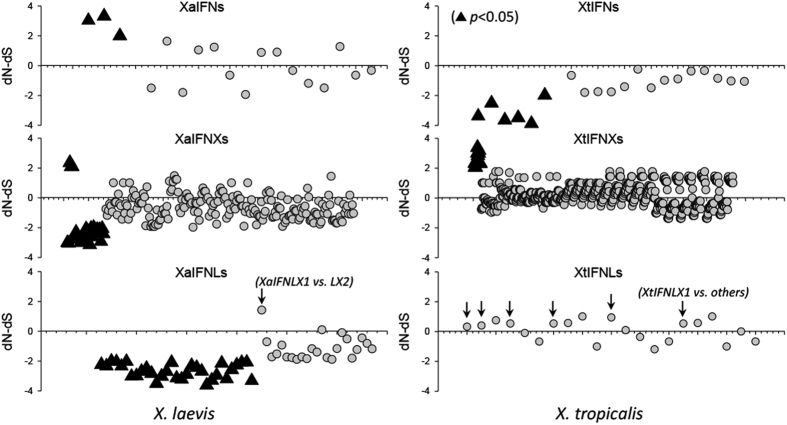
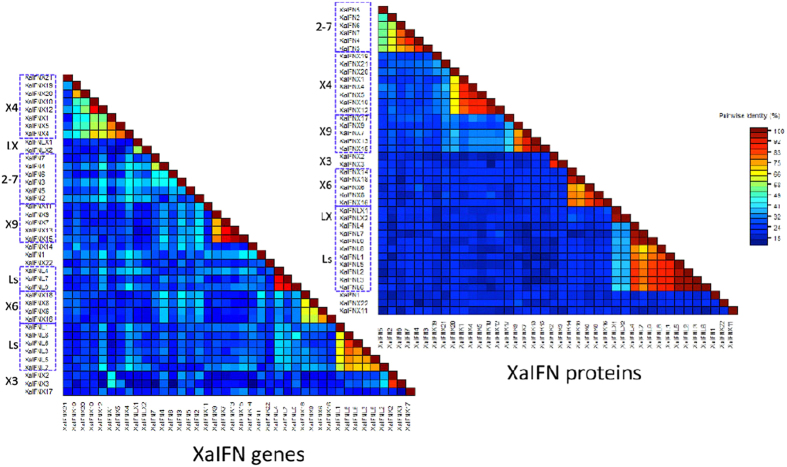
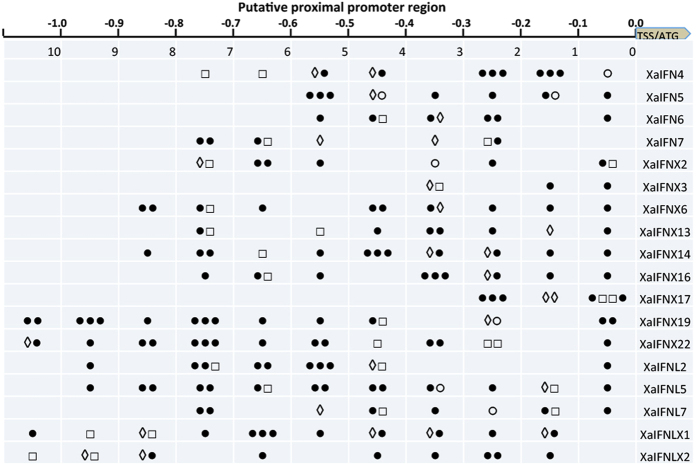
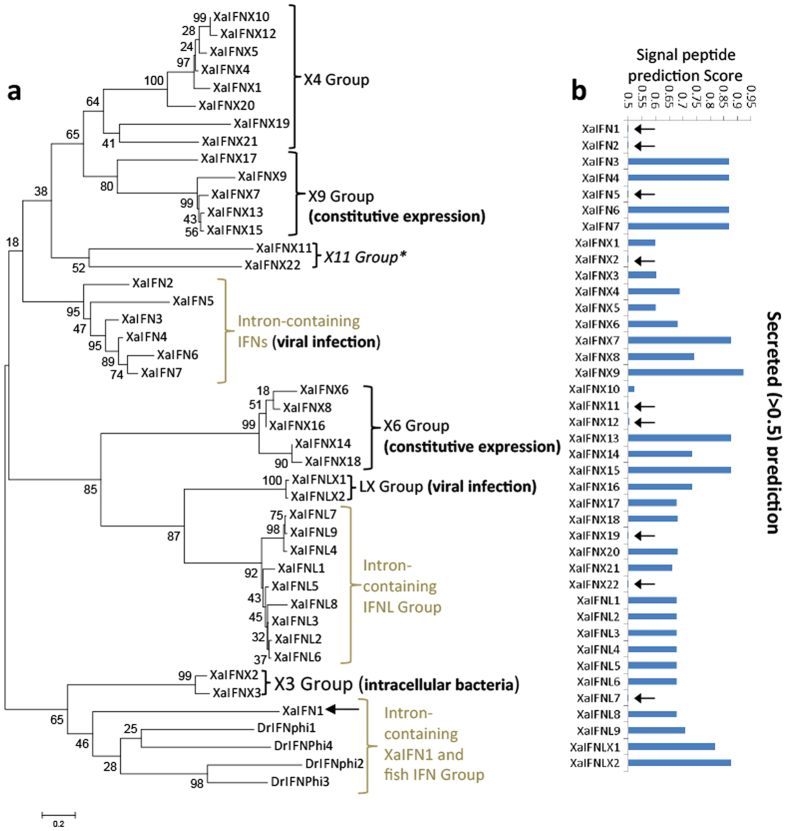
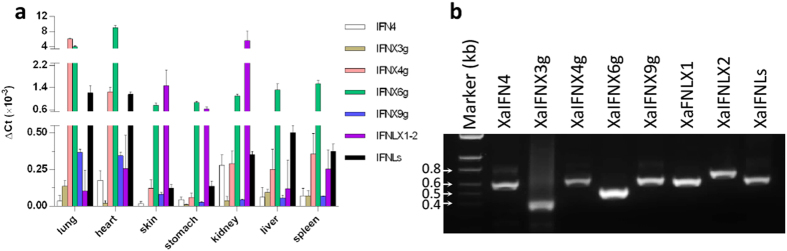
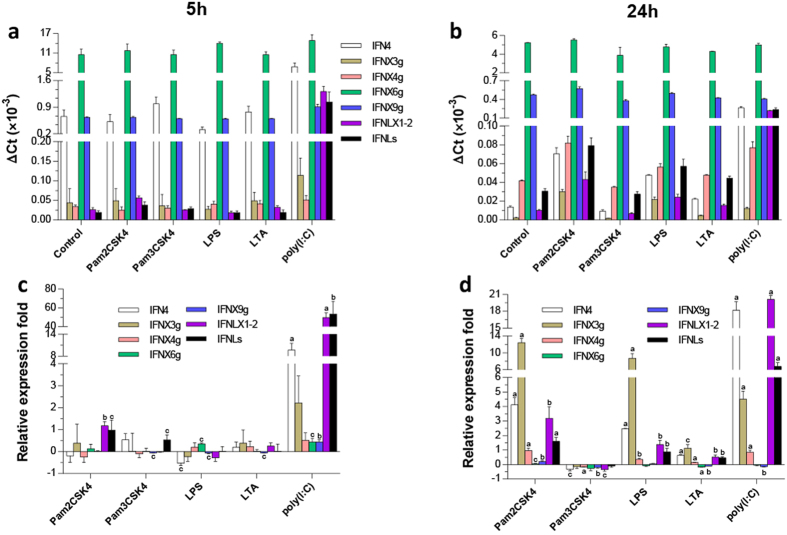
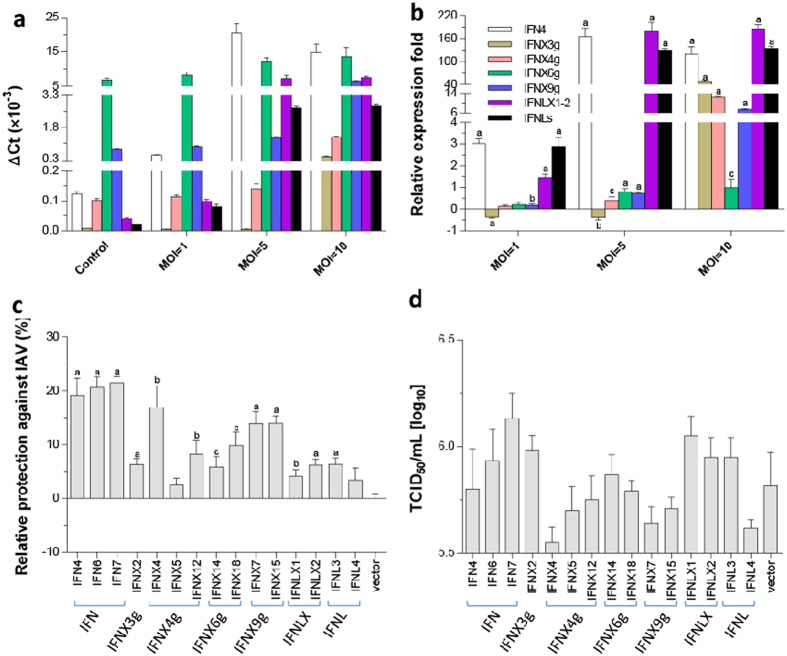
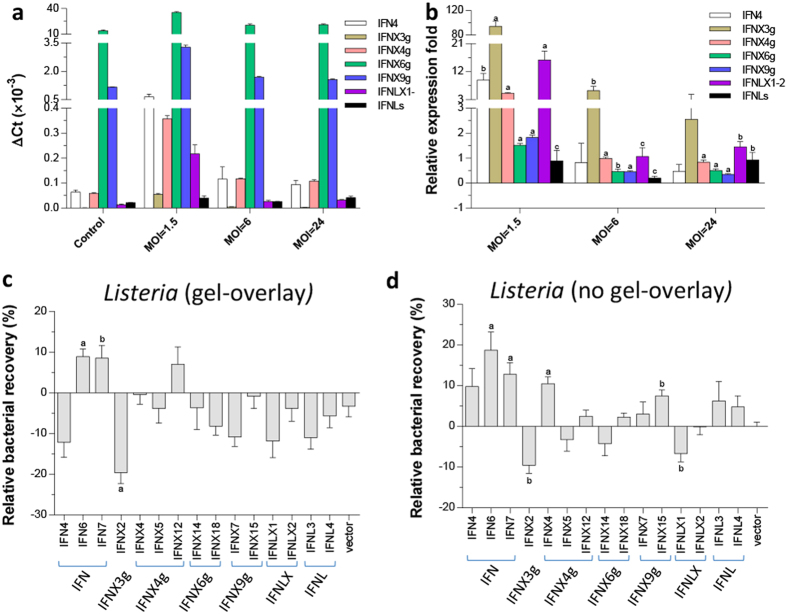
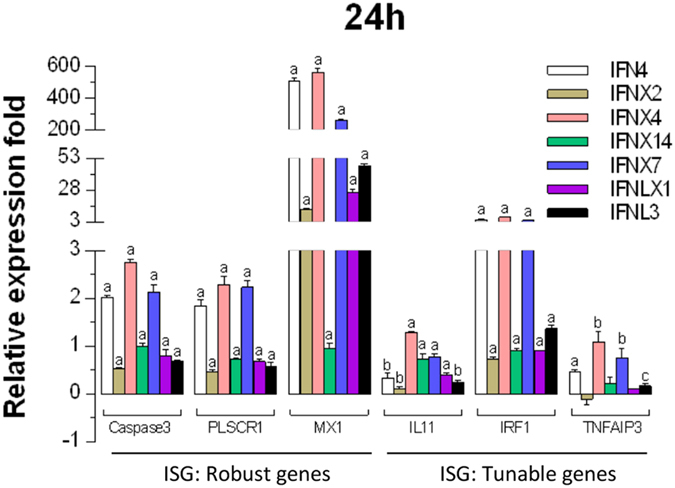
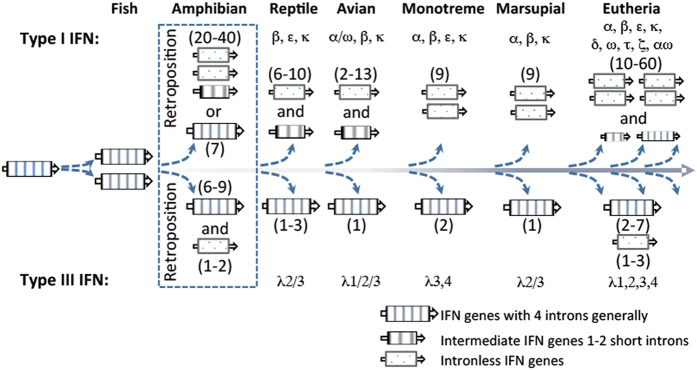
Interferons (IFNs) are key cytokines identified in vertebrates and evolutionary dominance of intronless IFN genes in amniotes is a signature event in IFN evolution. For the first time, we show that the emergence and expansion of intronless IFN genes is evident in amphibians, shown by 24-37 intronless IFN genes in each frog species. Amphibian IFNs represent a molecular complex more complicated than those in other vertebrate species, which revises the established model of IFN evolution to facilitate re-inspection of IFN molecular and functional diversity. We identified these intronless amphibian IFNs and their intron-containing progenitors, and functionally characterized constitutive and inductive expression and antimicrobial roles in infections caused by zoonotic pathogens, such as influenza viruses and Listeria monocytogenes. Amphibians, therefore, may serve as overlooked vectors/hosts for zoonotic pathogens, and the amphibian IFN system provides a model to study IFN evolution in molecular and functional diversity in coping with dramatic environmental changes during terrestrial adaption.
Figures
References
-
- Zitvogel L., Galluzzi L., Kepp O., Smyth M. J. & Kroemer G. Type I interferons in anticancer immunity. Nat. Rev. Immunol. 15, 405–414 (2015). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
