

Vascular thiol isomerases
- PMID: 27357699
- PMCID: PMC4990854
- DOI: 10.1182/blood-2016-04-636456
Vascular thiol isomerases
Abstract
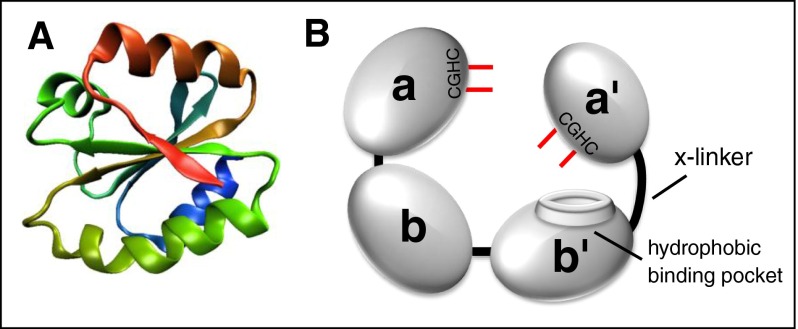
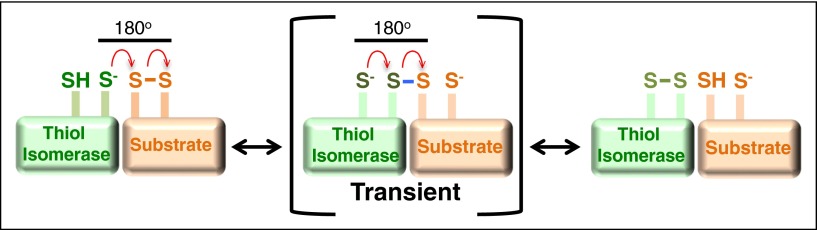
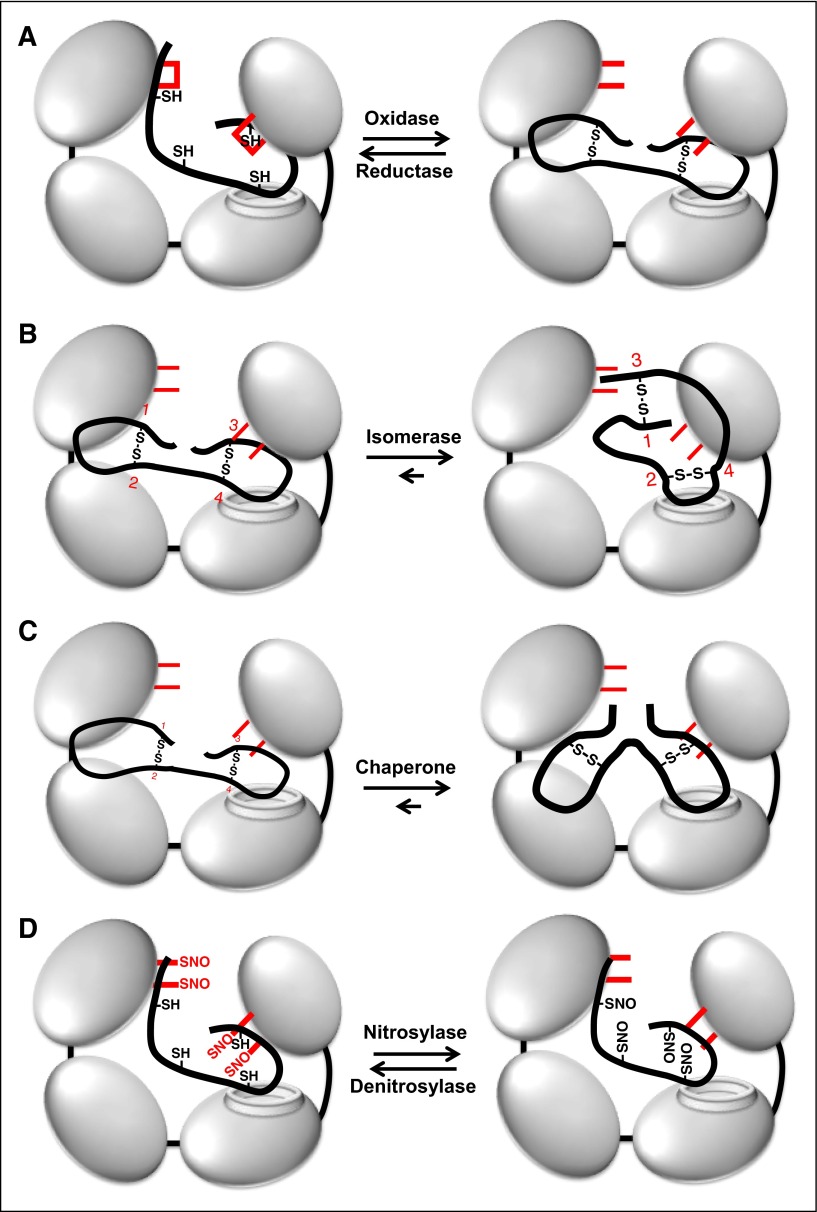
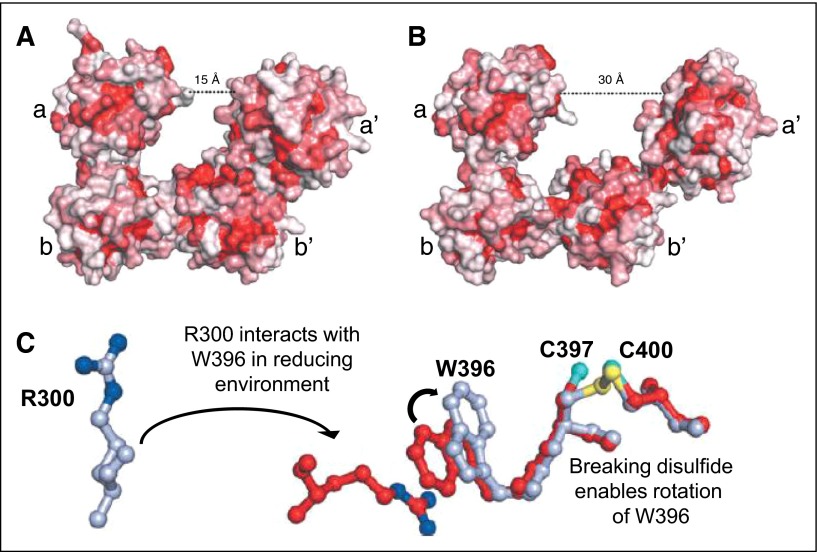
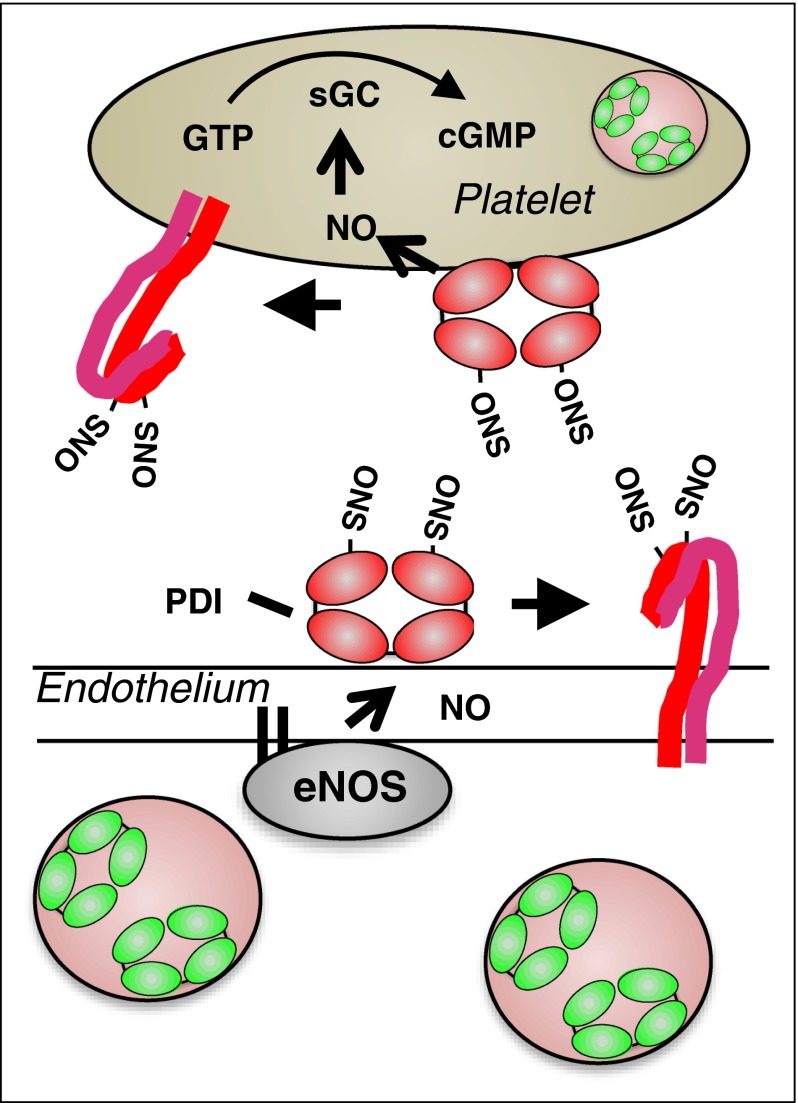
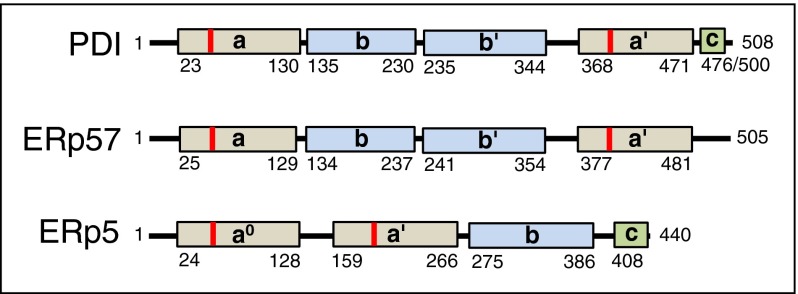
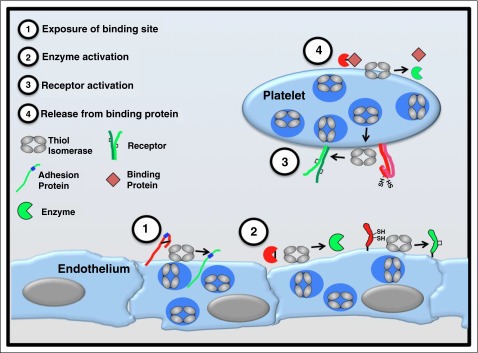
Thiol isomerases are multifunctional enzymes that influence protein structure via their oxidoreductase, isomerase, and chaperone activities. These enzymes localize at high concentrations in the endoplasmic reticulum of all eukaryotic cells where they serve an essential function in folding nascent proteins. However, thiol isomerases can escape endoplasmic retention and be secreted and localized on plasma membranes. Several thiol isomerases including protein disulfide isomerase, ERp57, and ERp5 are secreted by and localize to the membranes of platelets and endothelial cells. These vascular thiol isomerases are released following vessel injury and participate in thrombus formation. Although most of the activities of vascular thiol isomerases that contribute to thrombus formation are yet to be defined at the molecular level, allosteric disulfide bonds that are modified by thiol isomerases have been described in substrates such as αIIbβ3, αvβ3, GPIbα, tissue factor, and thrombospondin. Vascular thiol isomerases also act as redox sensors. They respond to the local redox environment and influence S-nitrosylation of surface proteins on platelets and endothelial cells. Despite our rudimentary understanding of the mechanisms by which thiol isomerases control vascular function, the clinical utility of targeting them in thrombotic disorders is already being explored in clinical trials.
© 2016 by The American Society of Hematology.
Figures
References
-
- Sela M, White FH, Jr, Anfinsen CB. Reductive cleavage of disulfide bridges in ribonuclease. Science. 1957;125(3250):691–692. - PubMed
-
- Dani HM, Fielder JA, Rabin BR. Degranulation and sex-specific regranulation of reticular membranes from rat liver as studied using a spectrophotometric assay of protein disulphide isomerase. FEBS Lett. 1976;65(3):377–382. - PubMed
-
- Goldberger RF, Epstein CJ, Anfinsen CB. Acceleration of reactivation of reduced bovine pancreatic ribonuclease by a microsomal system from rat liver. J Biol Chem. 1963;238:628–635. - PubMed
-
- Bulleid NJ, Freedman RB. Defective co-translational formation of disulphide bonds in protein disulphide-isomerase-deficient microsomes. Nature. 1988;335(6191):649–651. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
