

The Meiotic Recombination Activator PRDM9 Trimethylates Both H3K36 and H3K4 at Recombination Hotspots In Vivo
- PMID: 27362481
- PMCID: PMC4928815
- DOI: 10.1371/journal.pgen.1006146
The Meiotic Recombination Activator PRDM9 Trimethylates Both H3K36 and H3K4 at Recombination Hotspots In Vivo
Abstract
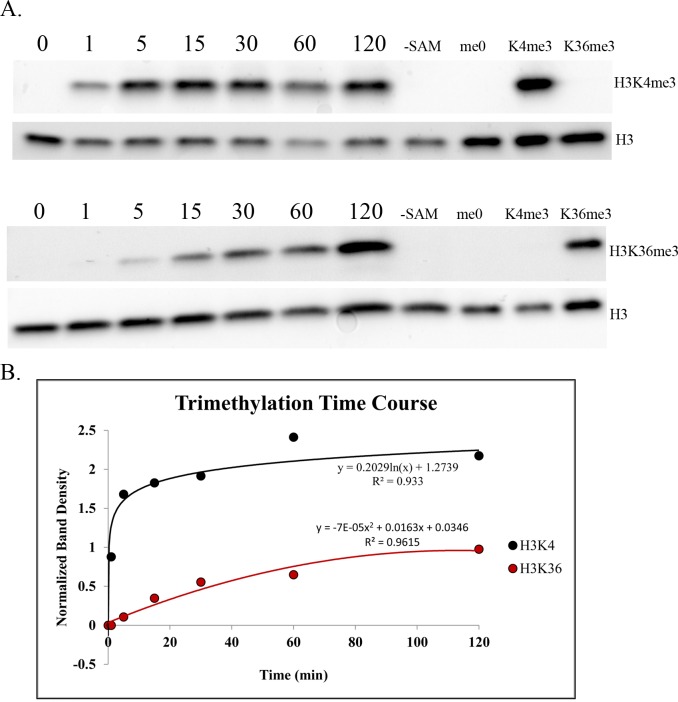
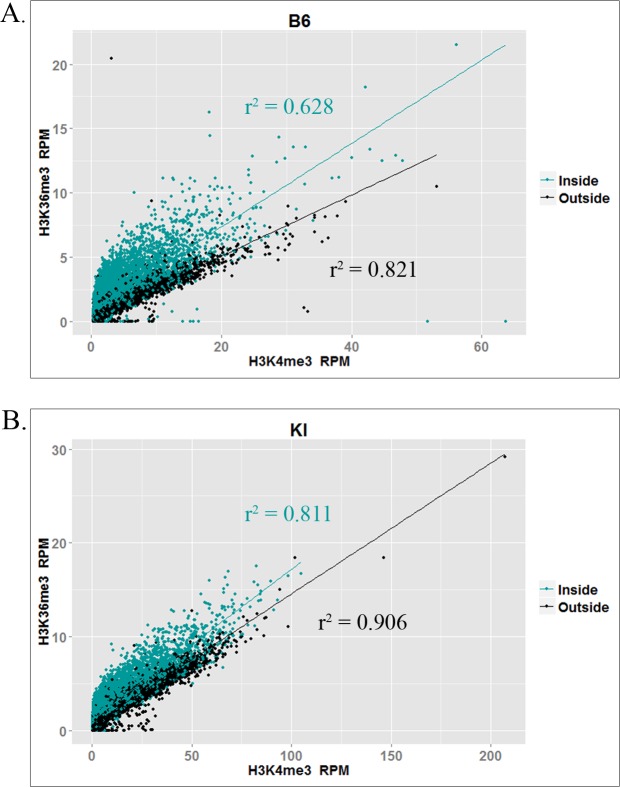
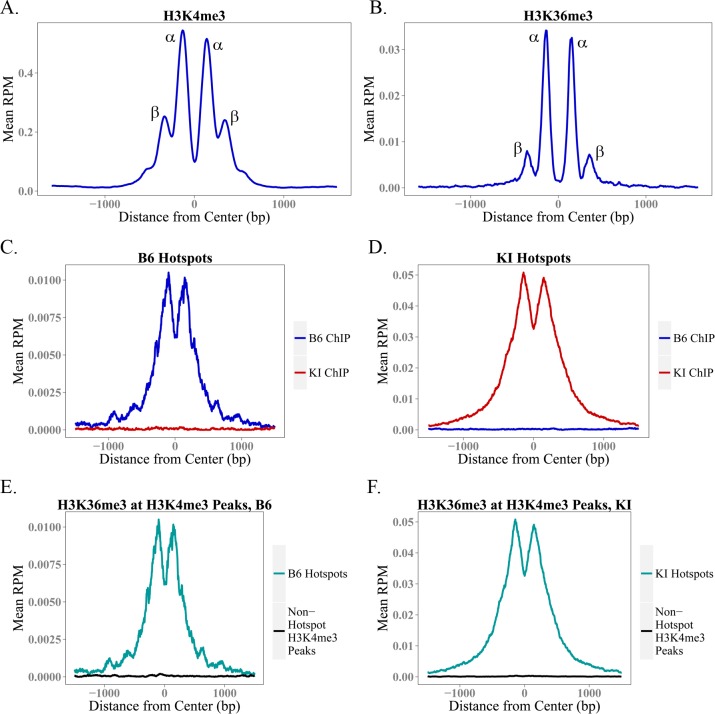
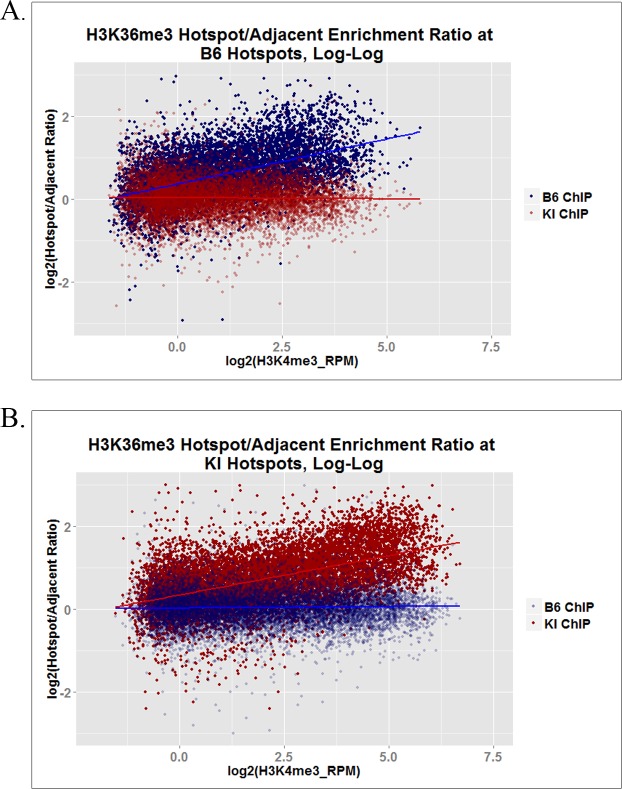
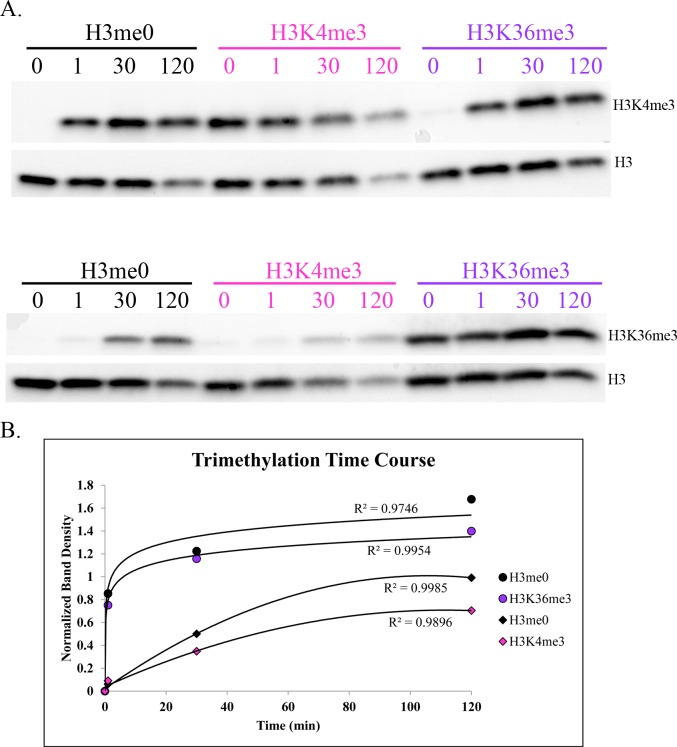
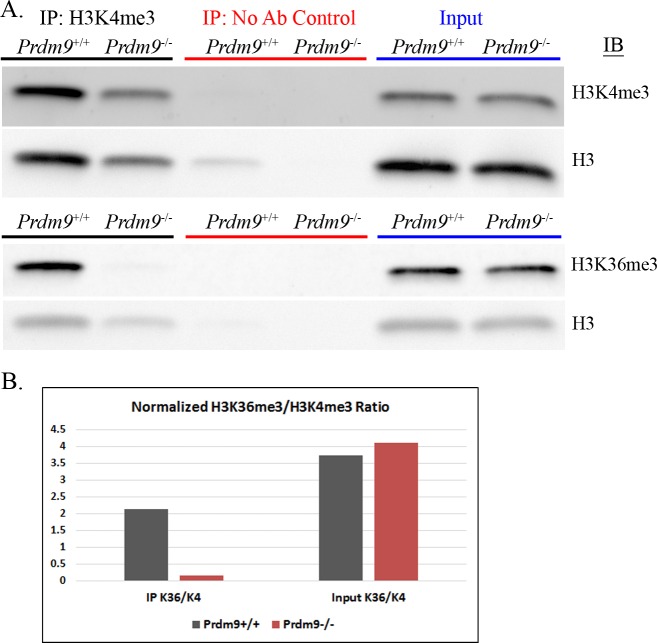
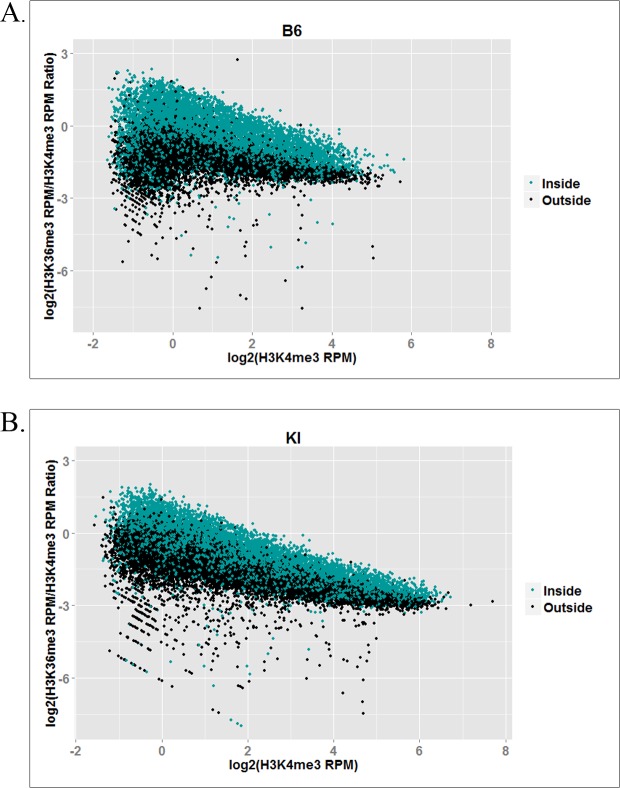
In many mammals, including humans and mice, the zinc finger histone methyltransferase PRDM9 performs the first step in meiotic recombination by specifying the locations of hotspots, the sites of genetic recombination. PRDM9 binds to DNA at hotspots through its zinc finger domain and activates recombination by trimethylating histone H3K4 on adjacent nucleosomes through its PR/SET domain. Recently, the isolated PR/SET domain of PRDM9 was shown capable of also trimethylating H3K36 in vitro, raising the question of whether this reaction occurs in vivo during meiosis, and if so, what its function might be. Here, we show that full-length PRDM9 does trimethylate H3K36 in vivo in mouse spermatocytes. Levels of H3K4me3 and H3K36me3 are highly correlated at hotspots, but mutually exclusive elsewhere. In vitro, we find that although PRDM9 trimethylates H3K36 much more slowly than it does H3K4, PRDM9 is capable of placing both marks on the same histone molecules. In accord with these results, we also show that PRDM9 can trimethylate both K4 and K36 on the same nucleosomes in vivo, but the ratio of K4me3/K36me3 is much higher for the pair of nucleosomes adjacent to the PRDM9 binding site compared to the next pair further away. Importantly, H3K4me3/H3K36me3-double-positive nucleosomes occur only in regions of recombination: hotspots and the pseudoautosomal (PAR) region of the sex chromosomes. These double-positive nucleosomes are dramatically reduced when PRDM9 is absent, showing that this signature is PRDM9-dependent at hotspots; the residual double-positive nucleosomes most likely come from the PRDM9-independent PAR. These results, together with the fact that PRDM9 is the only known mammalian histone methyltransferase with both H3K4 and H3K36 trimethylation activity, suggest that trimethylation of H3K36 plays an important role in the recombination process. Given the known requirement of H3K36me3 for double strand break repair by homologous recombination in somatic cells, we suggest that it may play the same role in meiosis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
