

Rickettsia australis Activates Inflammasome in Human and Murine Macrophages
- PMID: 27362650
- PMCID: PMC4928923
- DOI: 10.1371/journal.pone.0157231
Rickettsia australis Activates Inflammasome in Human and Murine Macrophages
Abstract
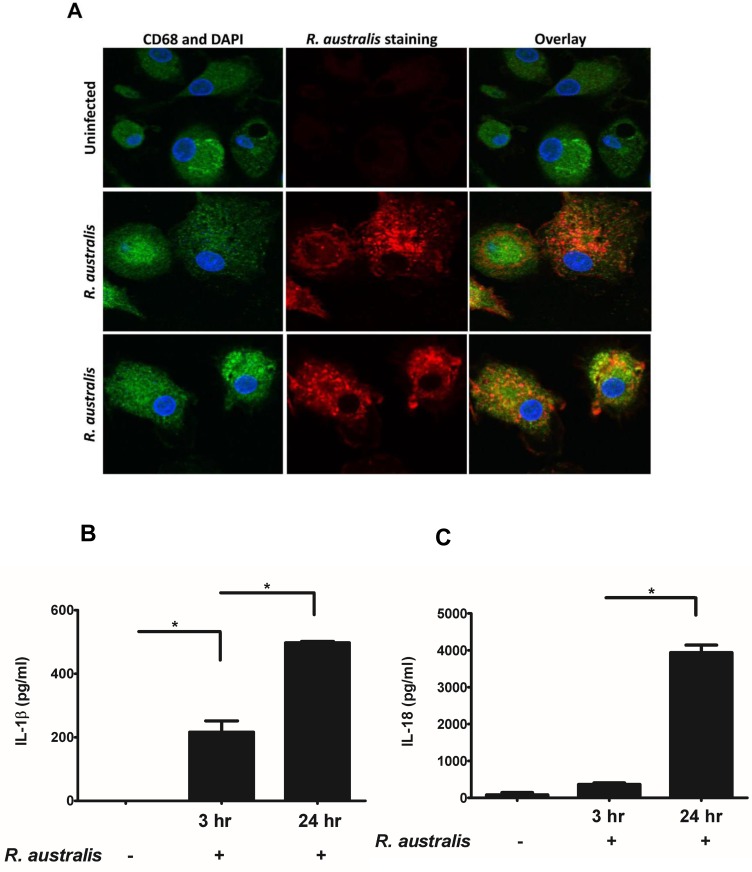
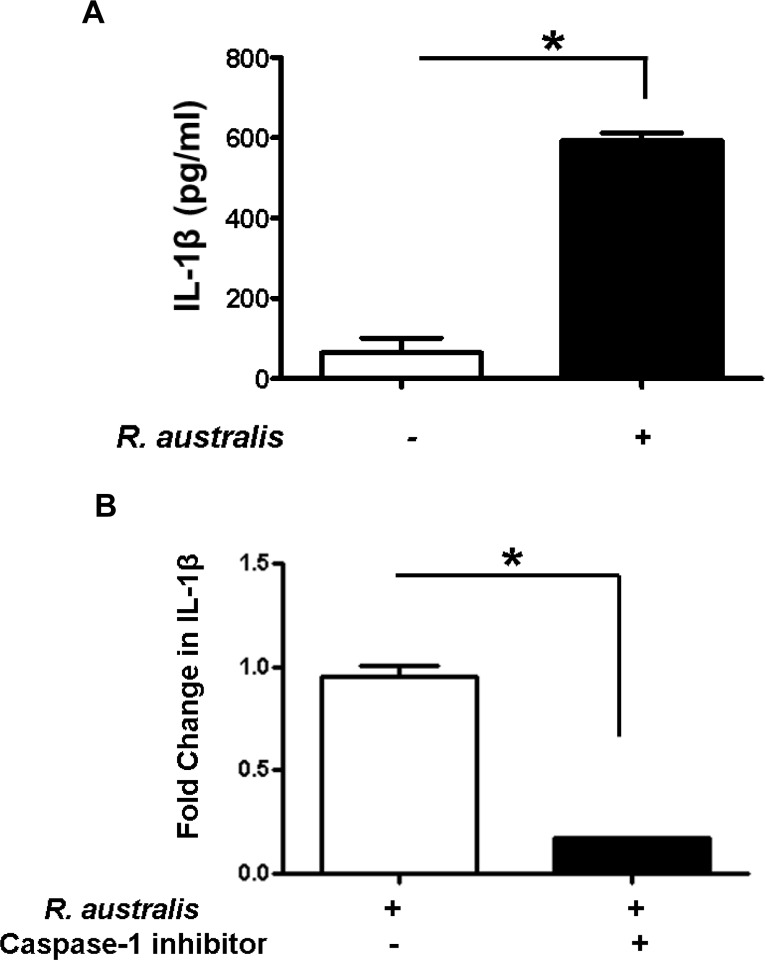
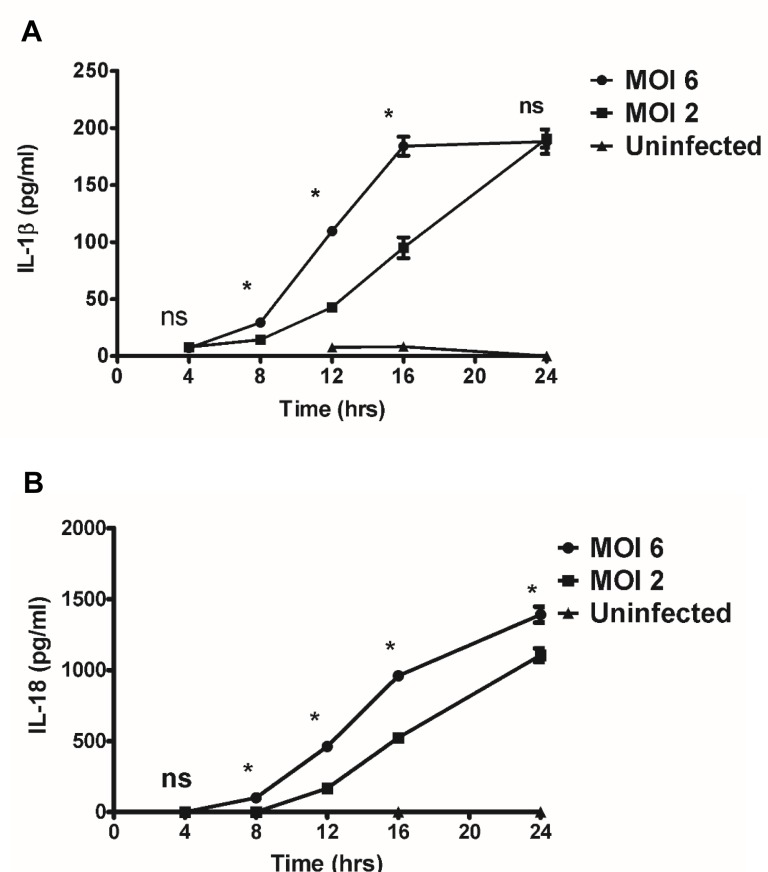
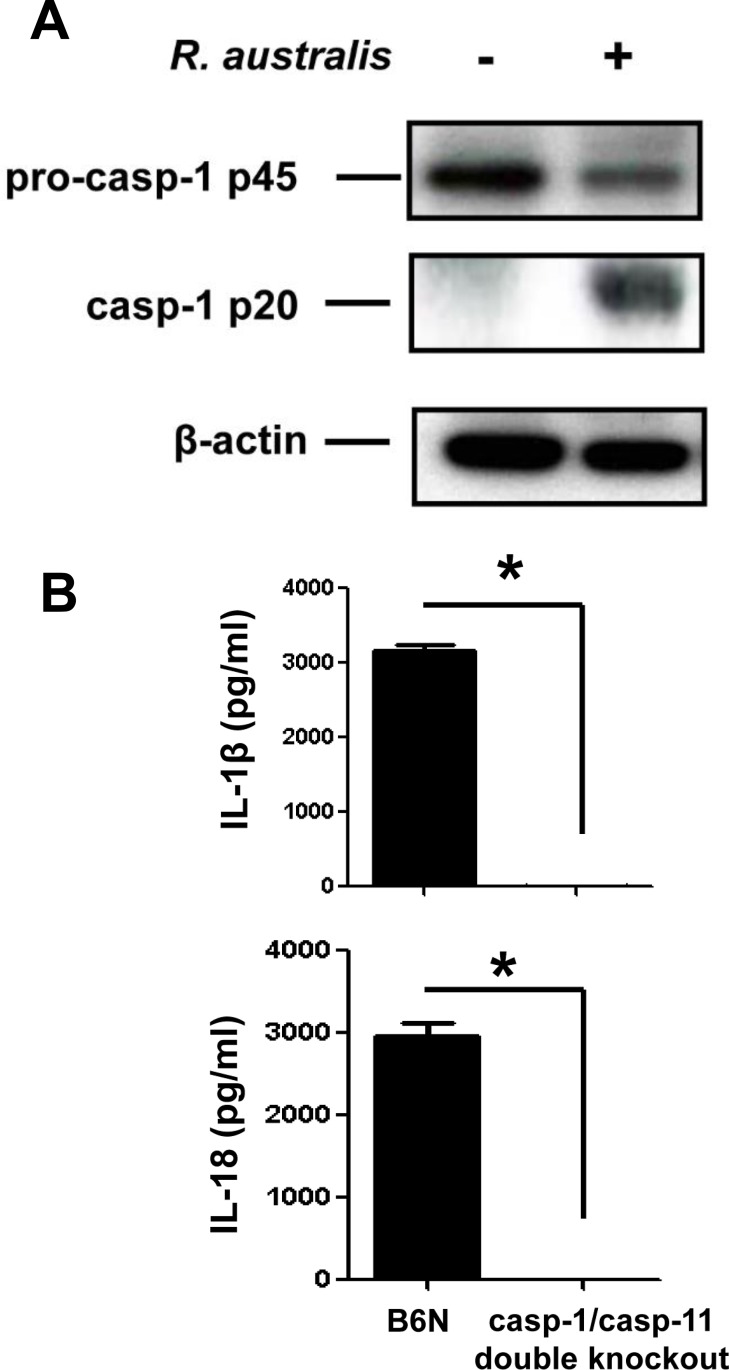
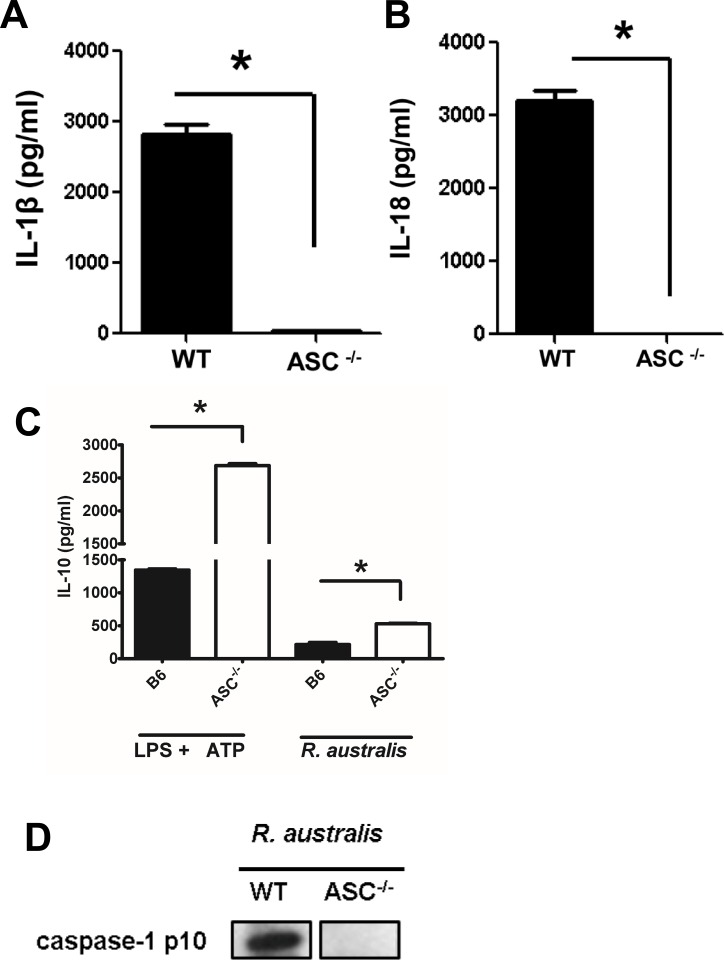
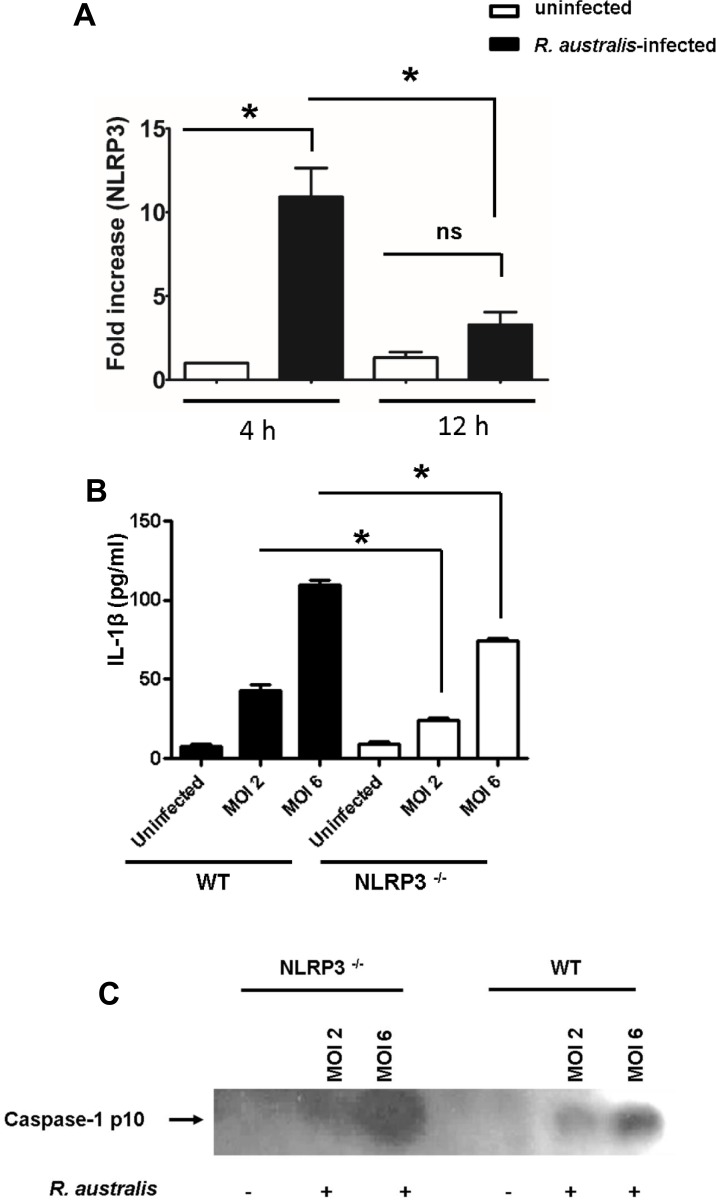
Rickettsiae actively escape from vacuoles and replicate free in the cytoplasm of host cells, where inflammasomes survey the invading pathogens. In the present study, we investigated the interactions of Rickettsia australis with the inflammasome in both mouse and human macrophages. R. australis induced a significant level of IL-1β secretion by human macrophages, which was significantly reduced upon treatment with an inhibitor of caspase-1 compared to untreated controls, suggesting caspase-1-dependent inflammasome activation. Rickettsia induced significant secretion of IL-1β and IL-18 in vitro by infected mouse bone marrow-derived macrophages (BMMs) as early as 8-12 h post infection (p.i.) in a dose-dependent manner. Secretion of these cytokines was accompanied by cleavage of caspase-1 and was completely abrogated in BMMs deficient in caspase-1/caspase-11 or apoptosis-associated speck-like protein containing a caspase activation and recruitment domain (ASC), suggesting that R. australis activate the ASC-dependent inflammasome. Interestingly, in response to the same quantity of rickettsiae, NLRP3-/- BMMs significantly reduced the secretion level of IL-1β compared to wild type (WT) controls, suggesting that NLRP3 inflammasome contributes to cytosolic recognition of R. australis in vitro. Rickettsial load in spleen, but not liver and lung, of R. australis-infected NLRP3-/- mice was significantly greater compared to WT mice. These data suggest that NLRP3 inflammasome plays a role in host control of bacteria in vivo in a tissue-specific manner. Taken together, our data, for the first time, illustrate the activation of ASC-dependent inflammasome by R. australis in macrophages in which NLRP3 is involved.
Conflict of interest statement
Figures
References
-
- Walker DH. Rickettsiae and rickettsial infections: the current state of knowledge. Clin Infect Dis. 2007; 45 Suppl 1:S39–44. - PubMed
-
- Feng HM, Popov VL, Walker DH. Depletion of gamma interferon and tumor necrosis factor alpha in mice with Rickettsia conorii-infected endothelium: impairment of rickettsicidal nitric oxide production resulting in fatal, overwhelming rickettsial disease. Infect Immun. 1994; 62(5):1952–60. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
