

Case Reports
doi: 10.3324/haematol.2016.145136.
Epub 2016 Jun 30.
Specific antibody deficiency and autoinflammatory disease extend the clinical and immunological spectrum of heterozygous NFKB1 loss-of-function mutations in humans
Affiliations
- PMID: 27365489
- PMCID: PMC5046658
- DOI: 10.3324/haematol.2016.145136
Item in Clipboard
Case Reports
Specific antibody deficiency and autoinflammatory disease extend the clinical and immunological spectrum of heterozygous NFKB1 loss-of-function mutations in humans
Haematologica.
2016 Oct.
No abstract available
Keywords: Fas; NFKB; antibody deficiency; autoimmune lymphoproliferative syndrome; autoinflammatory disease; immunodeficiency.
Figures
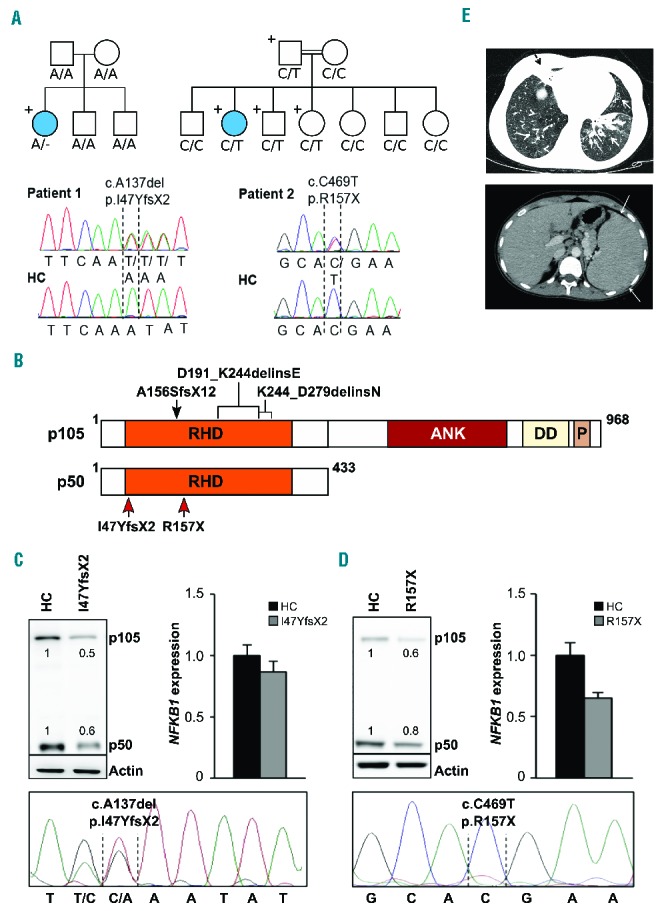
Two novel heterozygous NFKB1 mutations detected in two families
decrease the protein levels of p105 and p50. (A) Upper left: Whole exome
sequencing identified a heterozygous NFKB1 mutation
(A/−) in patient 1. The patient is the only carrier of the mutation in
the family pedigree (indicated by “+”) and the only diseased
family member (indicated by a filled circle). Lower left: Capillary sequencing
using genomic DNA confirmed an NFKB1 frameshift mutation
(c.A137del, p.I47YfsX2) in patient 1. Representative chromatograms of patient 1
and a healthy control (HC) are shown. Upper right: Patient 2 descended from
consanguineous parents and harbors an inherited heterozygous
NFKB1 mutation (C/T). The patient is the only diseased
family member. The father and two siblings carry the same mutation but are not
affected. Sanger sequencing of NFKB1 confirmed the heterozygous
missense mutation (c.C469T, p.R157X) in patient 2. Representative chromatograms
are shown. (B) Schematic drawing of the proteins p105 and p50 and their domains
which are both encoded by the NFKB1 gene. The mutations in the
Rel homology domain (RHD) identified in the two patients (red arrows) lead to
early truncation of both proteins. Previously reported heterozygous germline
mutations associated with CVID are indicated on top (black arrow and brackets).
ANK, ankyrin repeats; DD, death domain; P, PEST domain enriched for proline (P),
glutamic acid (E), serine (S), and threonine (T) residues. (C, D) Expression of
p105 and p50 proteins is decreased in the affected patients. (C) Primary T cells
of patient 1 and healthy controls were activated by phytohemagglutinin in the
presence of interleukin-2. Protein and RNA extracts were prepared. Western blot
analysis was carried out employing a specific p105/p50 antibody using
β-actin as a loading control (left panel). NFKB1 mRNA
expression was measured by real-time polymerase chain reaction (right panel).
The fold-change in cells of the patient compared to a representative healthy
control is shown, GAPDH and β-actin expression were used as internal
standards. Mean values of representative experiments performed in triplicates
and corresponding SDs are shown. Sanger sequencing using reverse transcribed
mRNA of the patient demonstrates the presence of mutated NFKB1
transcripts (lower panel). (D) Epstein-Barr virus-transformed B cells of patient
2 were used for protein and RNA extraction. Analysis of NFKB1 protein and RNA
expression was carried out as described in (C). Capillary sequencing of cDNA
from patient 2 failed to detect the NFKB1 mutation indicating
that the mutation leads to mRNA instability (lower panel). (E) Upper panel:
axial high resolution chest computer tomography image of patient 2 at the level
of lung bases demonstrating multiple areas of bronchiectasis (white arrows) and
consolidation with an atelectatic component surrounding bronchiectases in the
right middle lobe (black dashed arrow). Mosaic pattern of perfusion of the lung
parenchyma is noted, with multiple areas of low attenuation in the right low
lobe (arrowheads). Lower panel: axial computer tomography image of patient 2 at
the level of the upper abdomen demonstrating the enlarged spleen.
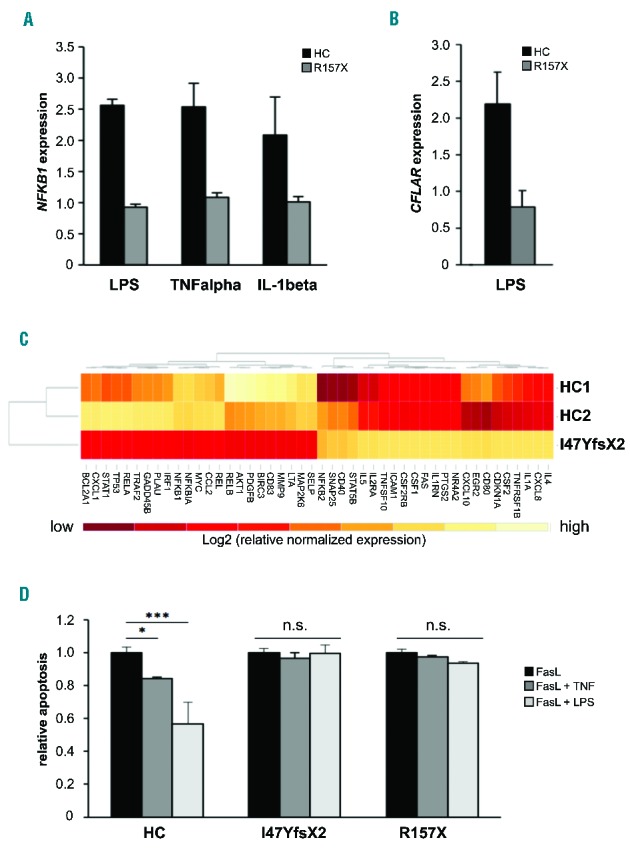
NF-κB-mediated signaling is affected in the patients. (A) Upregulation of
NFKB1 mRNA expression in response to NF-κB
activating stimuli is deficient in NFKB1-mutated cells.
Epstein-Barr virus (EBV)-immortalized B cells of patient 2 and of a
representative healthy control (HC) were treated for 16 h with LPS (5
μg/mL), tumor necrosis factor α (TNFα) (50 ng/mL) or
IL-1β (10 ng/mL). Fold change of NFKB1 mRNA expression
in untreated compared to treated samples was determined by real-time polymerase
chain reaction (PCR). The relative expression was normalized to the respective
untreated controls (=1). GAPDH and β-actin expression were used as
internal standards. Mean values of representative experiments performed in
triplicate and corresponding standard deviations are shown. (B) Expression of
the NF-κB target gene CFLAR (synonymous for cFLIP) is
not induced by LPS in the NFKB1-mutated patients’
cells. EBV-immortalized B cells of patient 2 and healthy control cells were
treated for 16 h with LPS (5 μg/mL). Fold change of mRNA expression of
CFLAR was determined by real-time PCR as described in Figure 1C. (C) Differential
expression of NF-κB target genes in NFKB1-mutated
primary T cells of patient 1 and two healthy wild-type controls. Baseline
expression of a panel of NF-κB target genes was analyzed by real-time
PCR using predesigned arrays (NF-κB signaling targets RT Profiler PCR arrays,
Qiagen, Hilden, Germany). Mean values of two independent assays are shown. High
gene expression is indicated in light color, low gene expression in dark red.
The analysis was supervised and results are shown scaled. (D) LPS does not
protect NFKB1 mutant primary T cells from apoptosis. Primary T
cells of patients 1 and 2 and healthy controls were activated by
phytohemagglutinin and IL2. Cells were treated with 100 ng/mL recombinant Fas
ligand to induce apoptosis in the presence or absence of 100 ng/mL LPS or 10
ng/mL TNFα. Apoptosis was determined by flow cytometric measurement of
phosphatidylserine exposure indicated by binding of annexin V-FITC. Dead cells
were detected with propidium iodide. Significance was tested using two-way ANoVA
(*P<0,05;
***P<0,001).
References
-
- Baeuerle PA, Henkel T. Function and activation of NF-kappa B in the immune system. Ann Rev Immunol. 1994;12:141–179. - PubMed
-
- Perkins ND. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat Rev Mol Cell Biol. 2007;8(1):49–62. - PubMed
-
- Pahl HL. Activators and target genes of Rel/NF-kappaB transcription factors. Oncogene. 1999;18(49):6853–6866. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
