

FcγR mediates TLR2- and Syk-dependent NLRP3 inflammasome activation by inactivated Francisella tularensis LVS immune complexes
- PMID: 27365531
- PMCID: PMC5110000
- DOI: 10.1189/jlb.2A1215-555RR
FcγR mediates TLR2- and Syk-dependent NLRP3 inflammasome activation by inactivated Francisella tularensis LVS immune complexes
Abstract
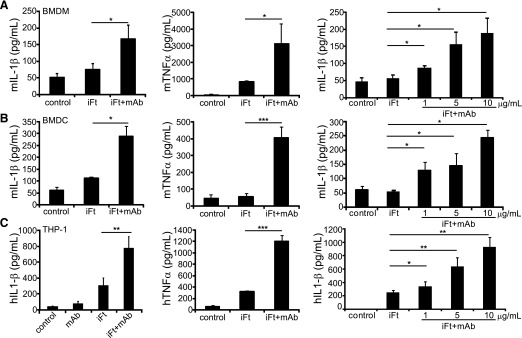
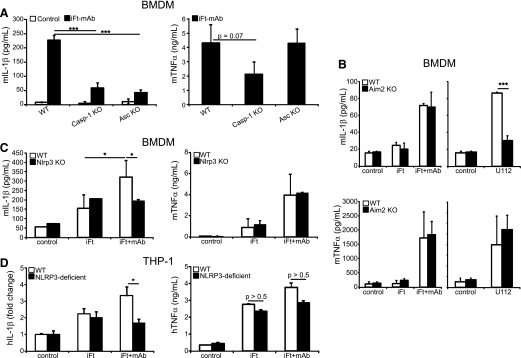
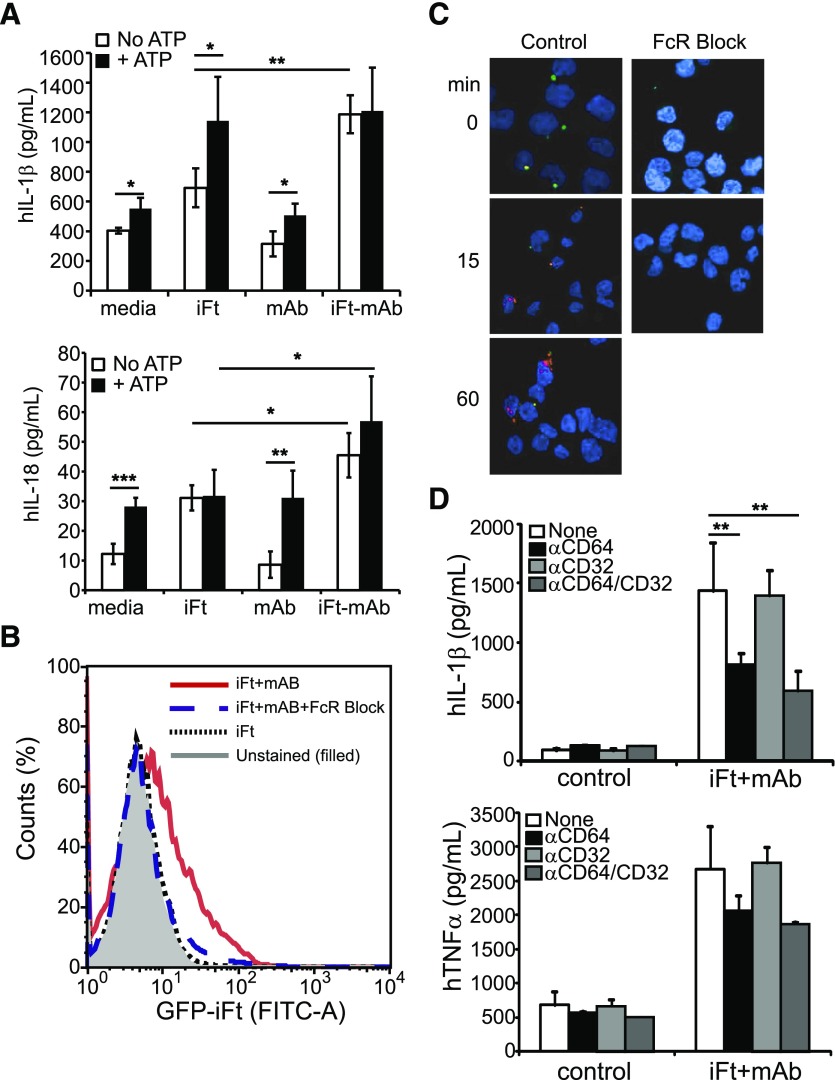
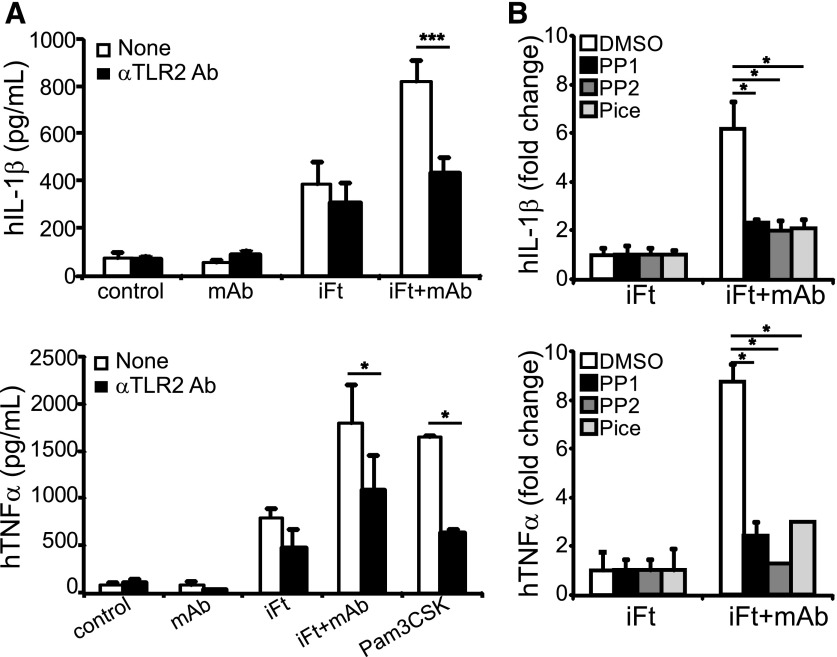
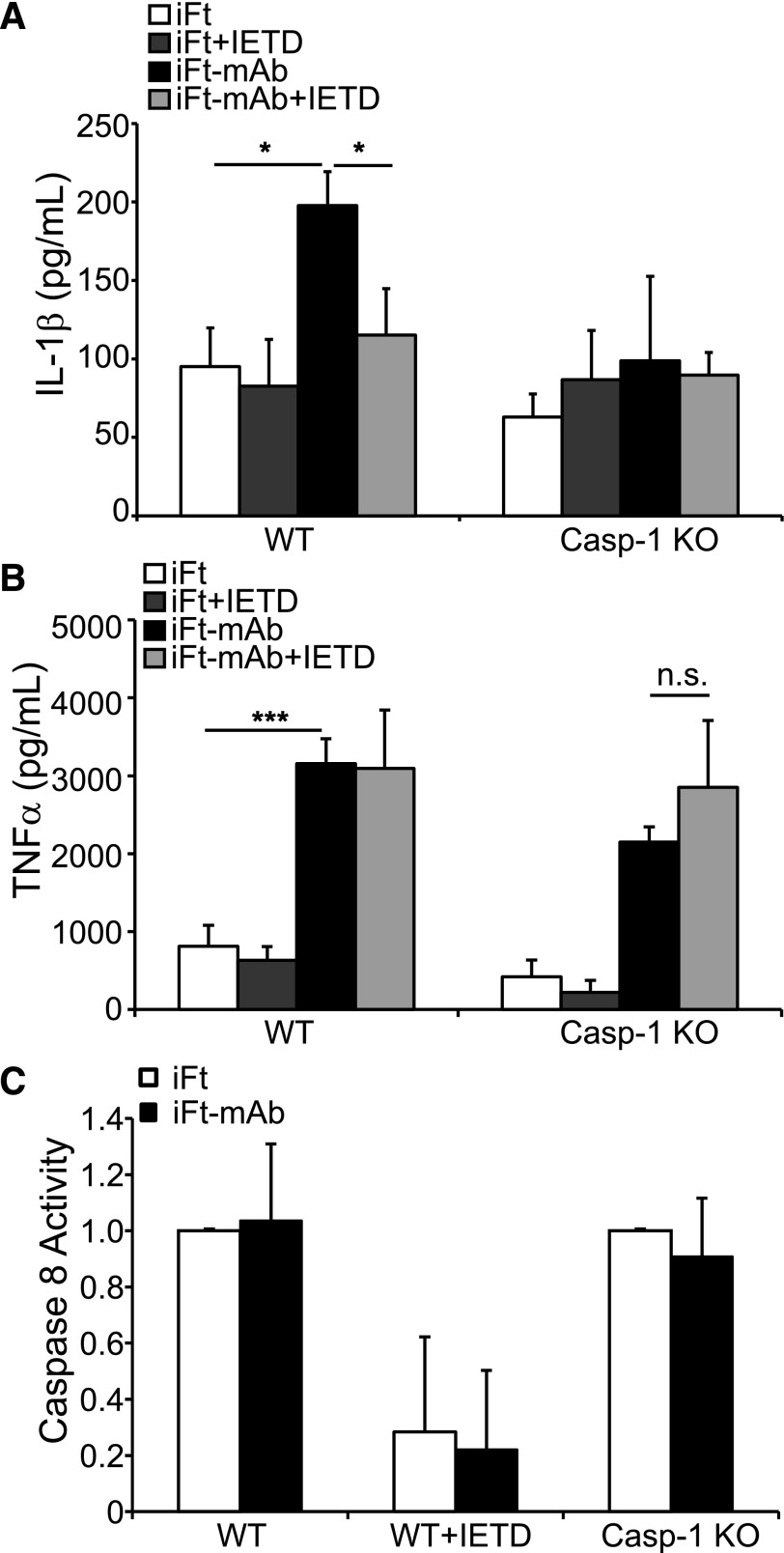
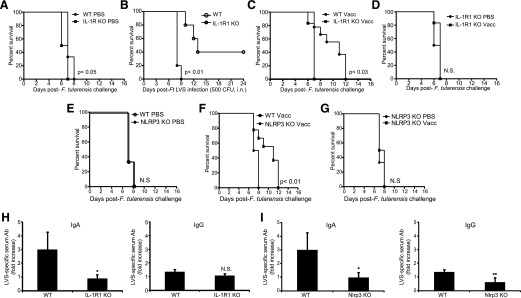
IgG (mAb)-opsonized, inactivated Francisella tularensis LVS (iFt-mAb) enhances TLR2-dependent IL-6 production by macrophages via Fcγ receptors (FcγR). In mice, vaccination with iFt-mAb provides IgA-dependent protection against lethal challenge with Ft LVS. Because inflammasome maturation of IL-1β is thought important for antibody-mediated immunity, we considered the possibility that iFt-mAb elicits an FcγR-dependent myeloid cell inflammasome response. Herein, we find that iFt-mAb enhances macrophage and dendritic cell IL-1β responses in a TLR2- and FcγR-dependent fashion. Although iFt-mAb complexes bind FcγR and are internalized, sensing of cytosolic DNA by absent in melanoma 2 (AIM2) is not required for the IL-1β response. In contrast, ASC, caspase-1, and NLR family pyrin domain-containing 3 (NLRP3) are indispensable. Further, FcγR-mediated spleen tyrosine kinase (Syk) signaling is required for this NLRP3-dependent IL-1β response, but the alternative IL-1β convertase caspase-8 is insufficient. Finally, iFt-mAb-vaccinated wild-type mice exhibit a significant delay in time to death, but IL-1R1- or Nlrp3-deficient mice vaccinated in this way are not protected and lack appreciable Francisella-specific antibodies. This study demonstrates that FcγR-mediated Syk activation leads to NLRP3 inflammasome-dependent IL-1β production in macrophages and suggests that an Nlrp3- and IL-1R-dependent process contributes to the IgA response important for protection against Ft LVS. These findings extend our understanding of cellular responses to inactivated pathogen-opsonized vaccine, establish FcγR-elicited Syk kinase-mediated NLRP3 inflammasome activation, and provide additional insight toward understanding crosstalk between TLR and FcγR signals.
Keywords: IL-1β; macrophage; vaccine.
© Society for Leukocyte Biology.
Figures
References
-
- Gross O., Poeck H., Bscheider M., Dostert C., Hannesschläger N., Endres S., Hartmann G., Tardivel A., Schweighoffer E., Tybulewicz V., Mocsai A., Tschopp J., Ruland J. (2009) Syk kinase signalling couples to the Nlrp3 inflammasome for anti-fungal host defence. Nature 459, 433–436. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
