

The zinc fingers of YY1 bind single-stranded RNA with low sequence specificity
- PMID: 27369384
- PMCID: PMC5100589
- DOI: 10.1093/nar/gkw590
The zinc fingers of YY1 bind single-stranded RNA with low sequence specificity
Abstract
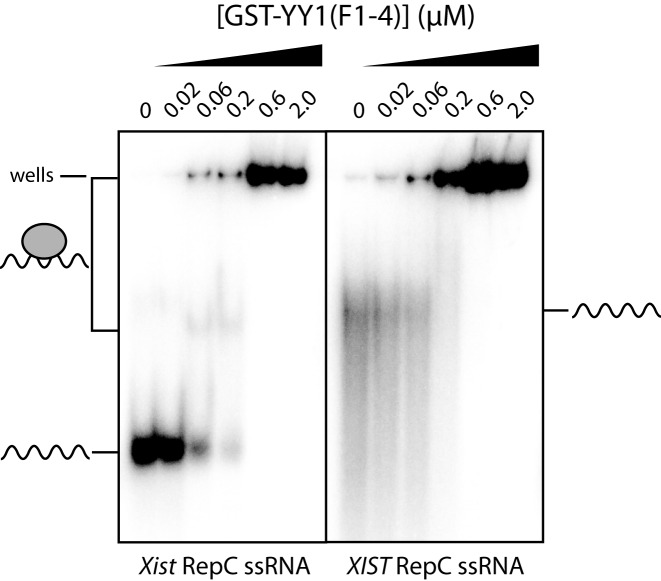
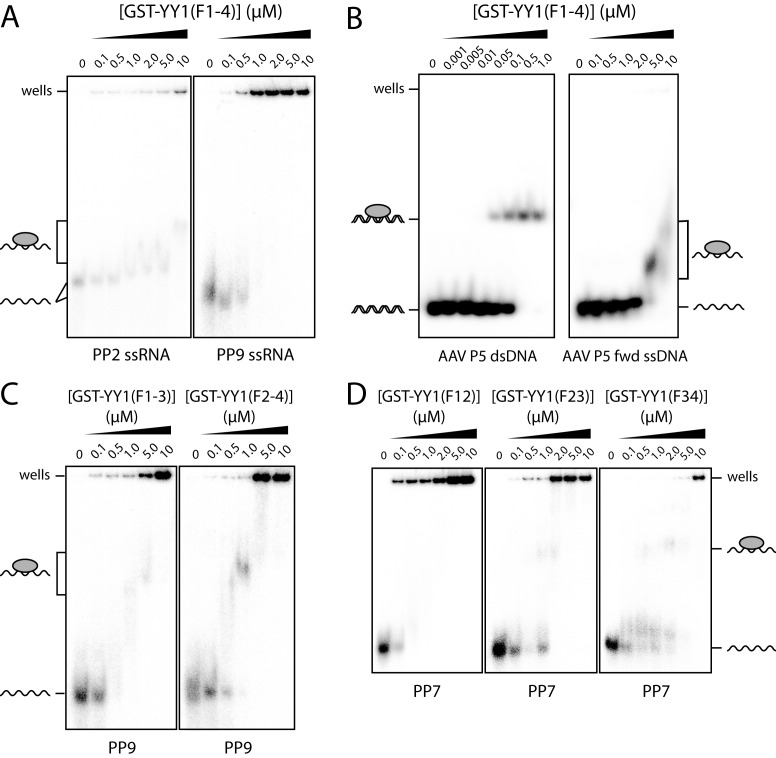
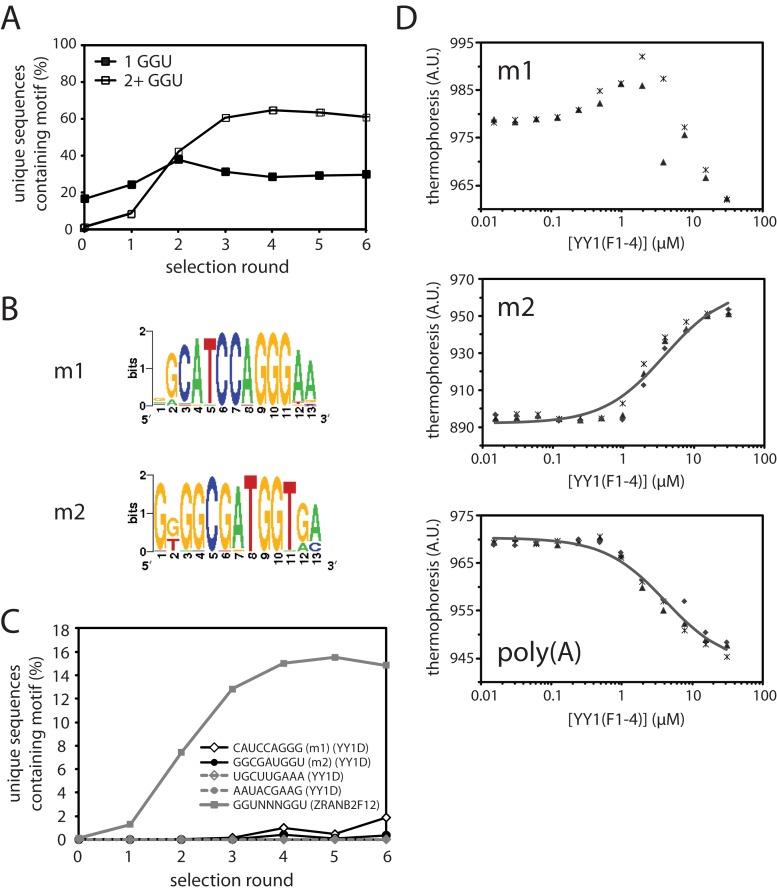
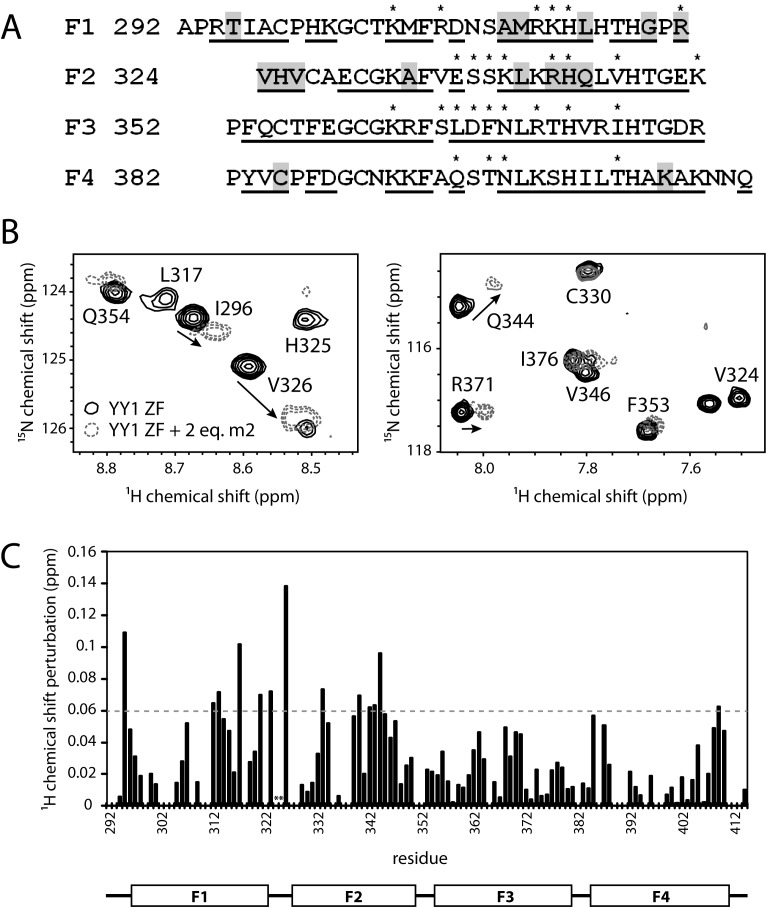
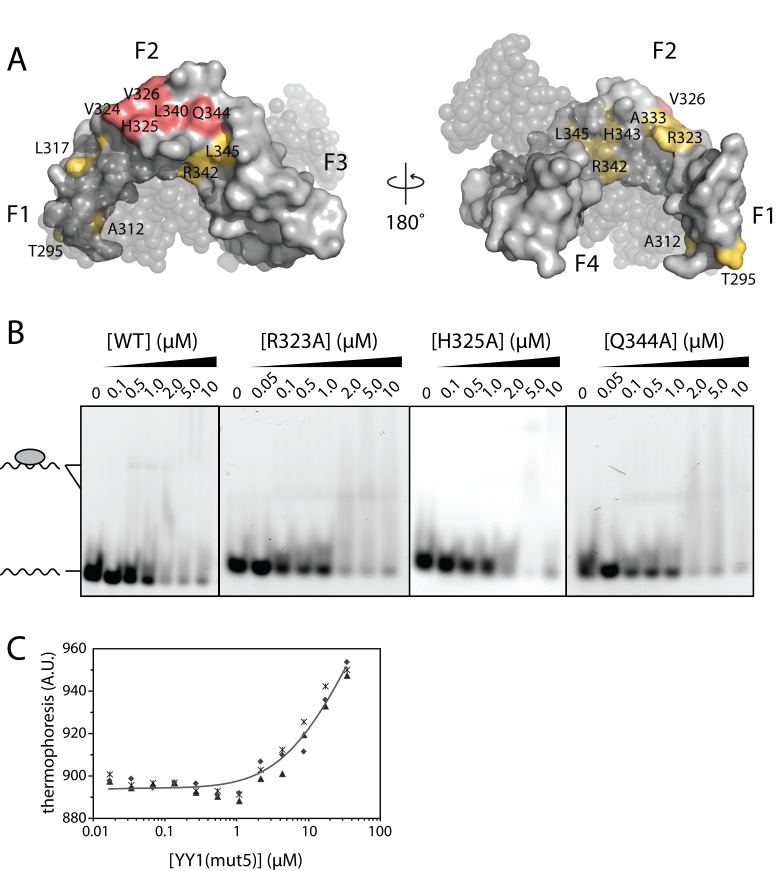
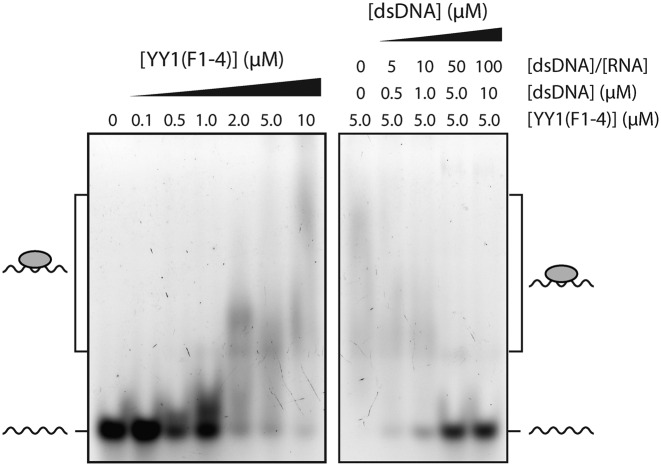
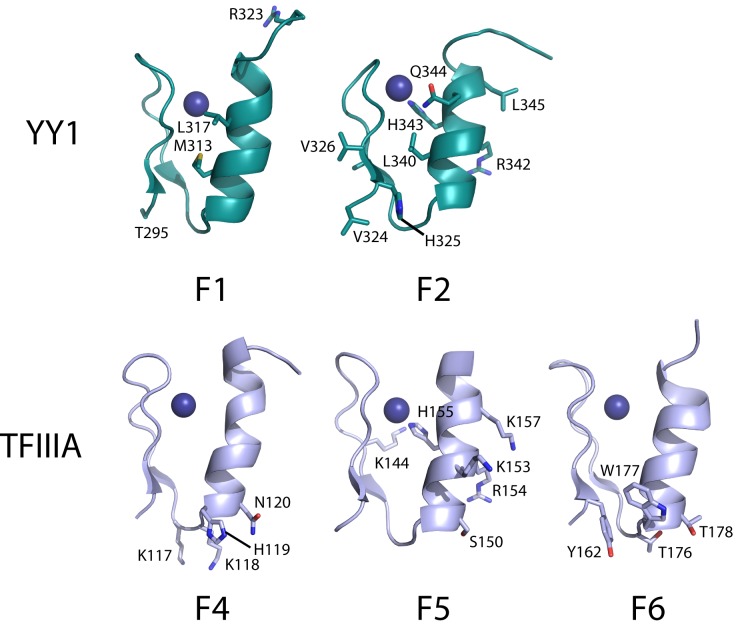
Classical zinc fingers (ZFs) are traditionally considered to act as sequence-specific DNA-binding domains. More recently, classical ZFs have been recognised as potential RNA-binding modules, raising the intriguing possibility that classical-ZF transcription factors are involved in post-transcriptional gene regulation via direct RNA binding. To date, however, only one classical ZF-RNA complex, that involving TFIIIA, has been structurally characterised. Yin Yang-1 (YY1) is a multi-functional transcription factor involved in many regulatory processes, and binds DNA via four classical ZFs. Recent evidence suggests that YY1 also interacts with RNA, but the molecular nature of the interaction remains unknown. In the present work, we directly assess the ability of YY1 to bind RNA using in vitro assays. Systematic Evolution of Ligands by EXponential enrichment (SELEX) was used to identify preferred RNA sequences bound by the YY1 ZFs from a randomised library over multiple rounds of selection. However, a strong motif was not consistently recovered, suggesting that the RNA sequence selectivity of these domains is modest. YY1 ZF residues involved in binding to single-stranded RNA were identified by NMR spectroscopy and found to be largely distinct from the set of residues involved in DNA binding, suggesting that interactions between YY1 and ssRNA constitute a separate mode of nucleic acid binding. Our data are consistent with recent reports that YY1 can bind to RNA in a low-specificity, yet physiologically relevant manner.
© The Author(s) 2016. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Functional analysis of YY1 zinc fingers through cysteine mutagenesis.FEBS Lett. 2019 Jun;593(12):1392-1402. doi: 10.1002/1873-3468.13431. Epub 2019 May 24. FEBS Lett. 2019. PMID: 31127623
-
Structural metal sites in nonclassical zinc finger proteins involved in transcriptional and translational regulation.Acc Chem Res. 2014 Aug 19;47(8):2643-50. doi: 10.1021/ar500182d. Epub 2014 Aug 6. Acc Chem Res. 2014. PMID: 25098749
-
Physical Interaction of Human Yin Yang 1 Protein with DNA.Crit Rev Oncog. 2017;22(1-2):75-97. doi: 10.1615/CritRevOncog.2017020976. Crit Rev Oncog. 2017. PMID: 29604938 Review.
-
Identification and characterization of four novel peptide motifs that recognize distinct regions of the transcription factor CP2.FEBS J. 2005 Mar;272(5):1265-77. doi: 10.1111/j.1742-4658.2005.04564.x. FEBS J. 2005. PMID: 15720400
-
Two-timing zinc finger transcription factors liaising with RNA.Trends Biochem Sci. 2012 May;37(5):199-205. doi: 10.1016/j.tibs.2012.02.001. Epub 2012 Mar 7. Trends Biochem Sci. 2012. PMID: 22405571 Review.
Cited by
-
Interactome of vertebrate GAF/ThPOK reveals its diverse functions in gene regulation and DNA repair.J Biosci. 2020;45:38. J Biosci. 2020. PMID: 32098917
-
Yin Yang 1 is a potent activator of human T lymphotropic virus type 1 LTR-driven gene expression via RNA binding.Proc Natl Acad Sci U S A. 2020 Aug 4;117(31):18701-18710. doi: 10.1073/pnas.2005726117. Epub 2020 Jul 20. Proc Natl Acad Sci U S A. 2020. PMID: 32690679 Free PMC article.
-
How do lncRNAs regulate transcription?Sci Adv. 2017 Sep 27;3(9):eaao2110. doi: 10.1126/sciadv.aao2110. eCollection 2017 Sep. Sci Adv. 2017. PMID: 28959731 Free PMC article. Review.
-
Loss of Xist RNA from the inactive X during B cell development is restored in a dynamic YY1-dependent two-step process in activated B cells.PLoS Genet. 2017 Oct 9;13(10):e1007050. doi: 10.1371/journal.pgen.1007050. eCollection 2017 Oct. PLoS Genet. 2017. PMID: 28991910 Free PMC article.
-
Deciphering the DNA code for the function of the Drosophila polydactyl zinc finger protein Suppressor of Hairy-wing.Nucleic Acids Res. 2017 May 5;45(8):4463-4478. doi: 10.1093/nar/gkx040. Nucleic Acids Res. 2017. PMID: 28158673 Free PMC article.
References
-
- Razin S.V., Borunova V.V., Maksimenko O.G., Kantidze O.L. Cys2His2 zinc finger protein family: Classification, functions, and major members. Biochemistry-Moscow. 2012;77:217–226. - PubMed
-
- Brayer K., Segal D. Keep your fingers Off My DNA: protein–protein interactions mediated by C2H2 zinc finger domains. Cell Biochem. Biophys. 2008;50:111–131. - PubMed
-
- Lu D., Alexandra Searles M., Klug A. Crystal structure of a zinc-finger-RNA complex reveals two modes of molecular recognition. Nature. 2003;426:96–100. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
