

BRCA1/FANCD2/BRG1-Driven DNA Repair Stabilizes the Differentiation State of Human Mammary Epithelial Cells
- PMID: 27373334
- PMCID: PMC4982517
- DOI: 10.1016/j.molcel.2016.05.038
BRCA1/FANCD2/BRG1-Driven DNA Repair Stabilizes the Differentiation State of Human Mammary Epithelial Cells
Abstract
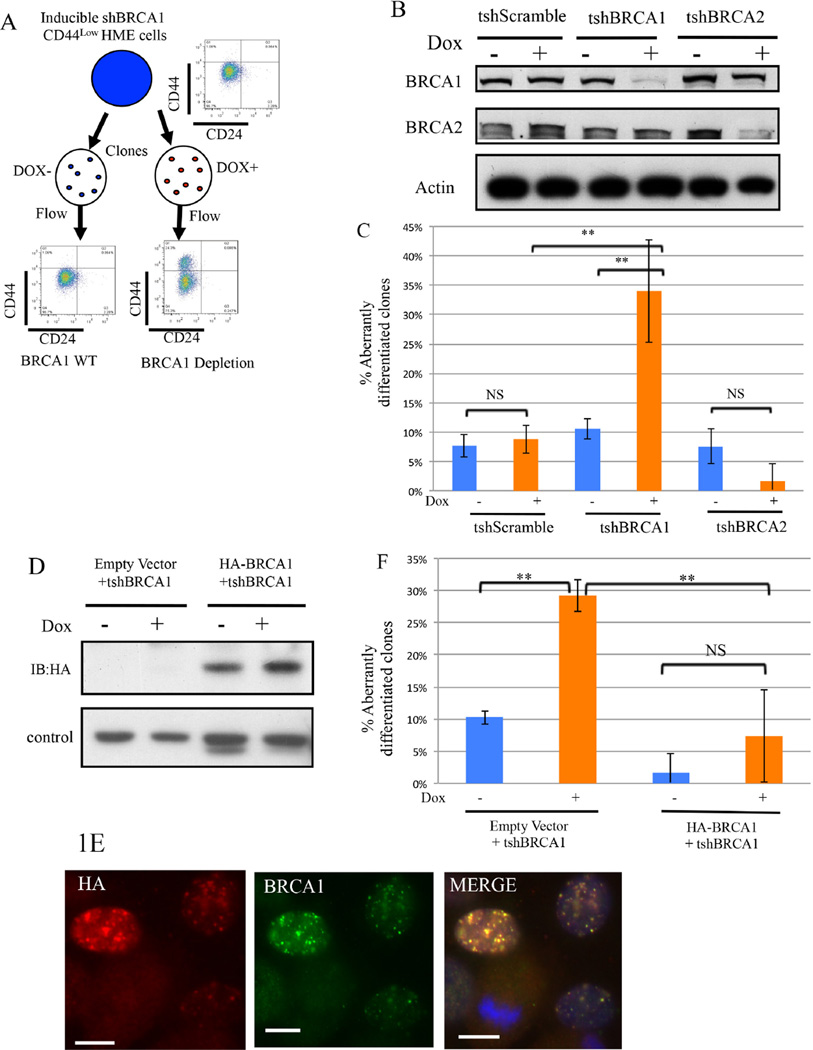
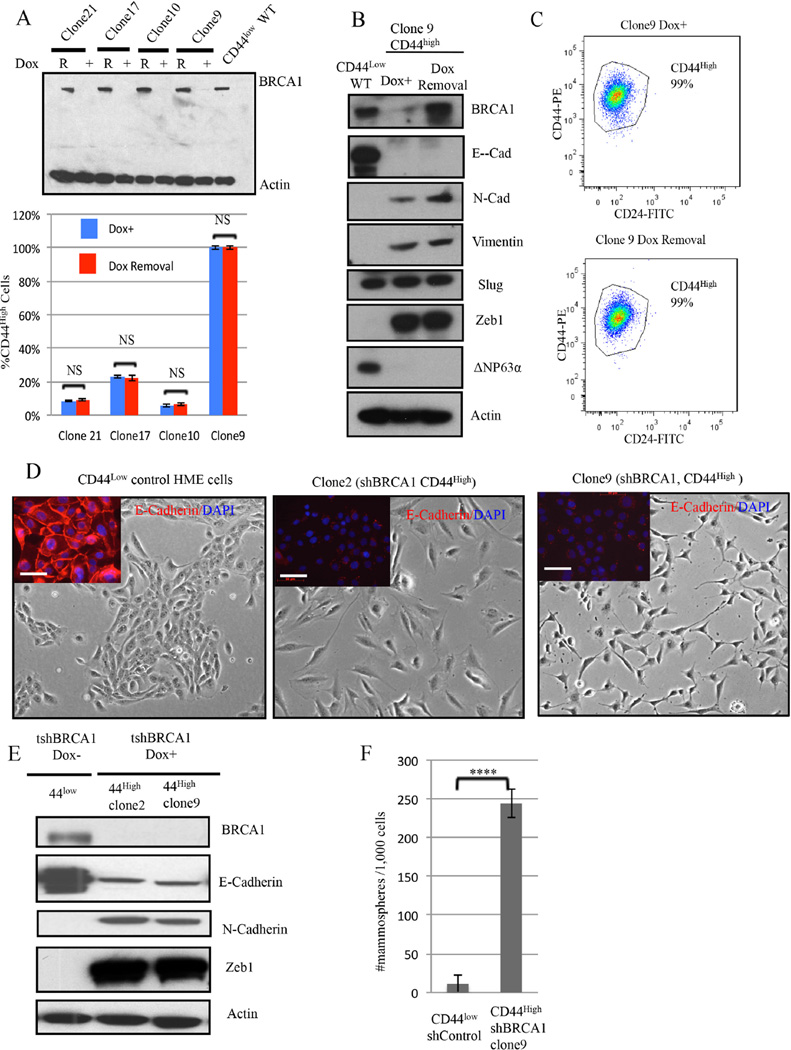
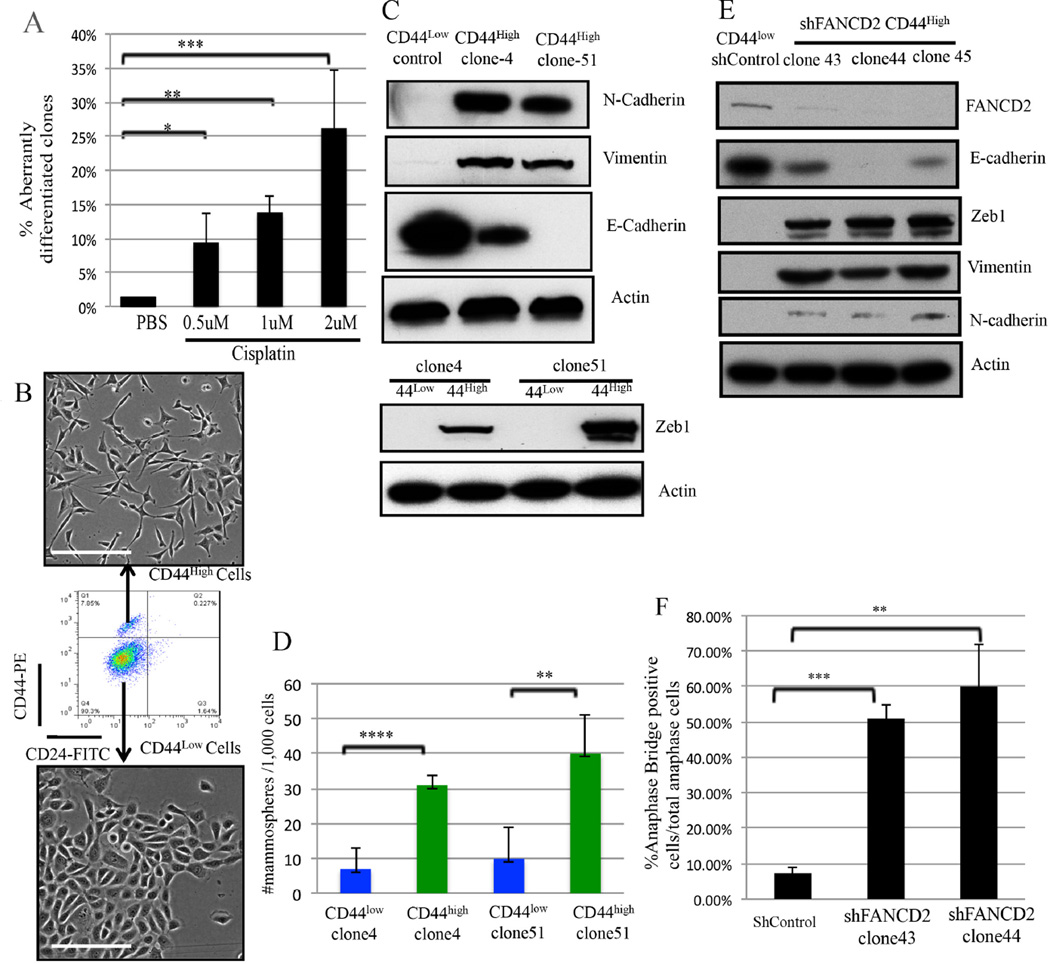
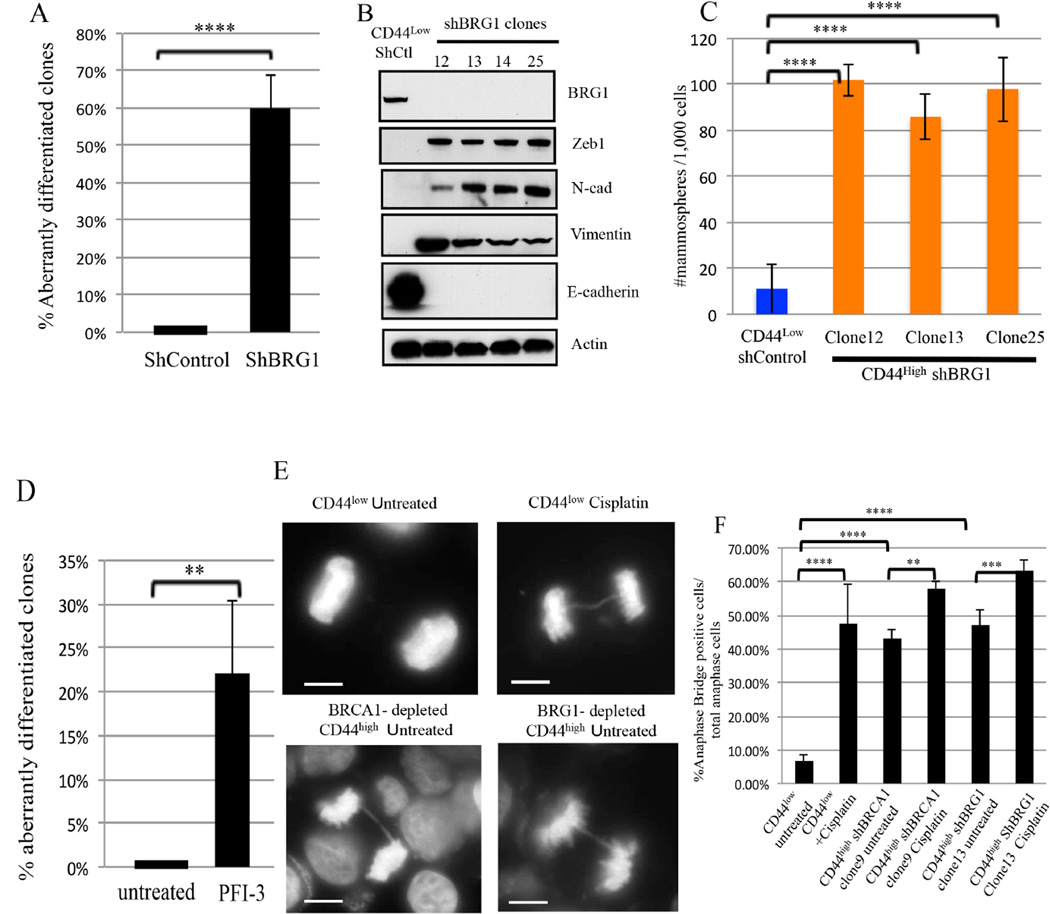
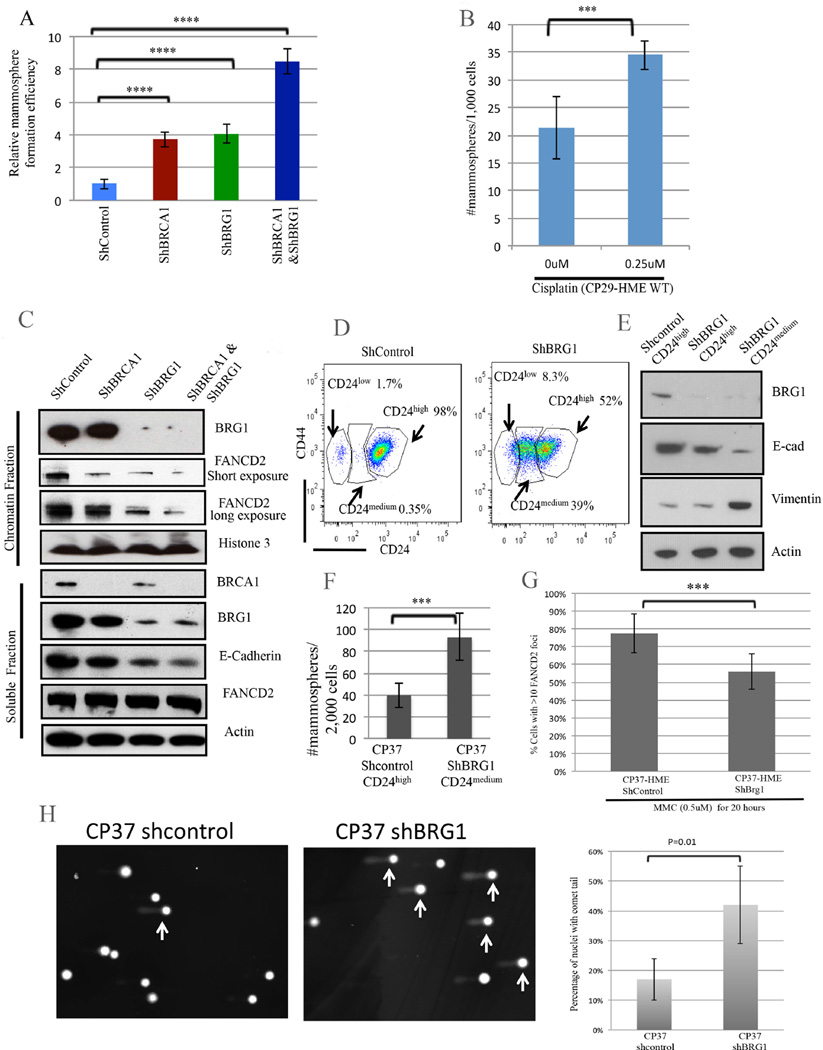
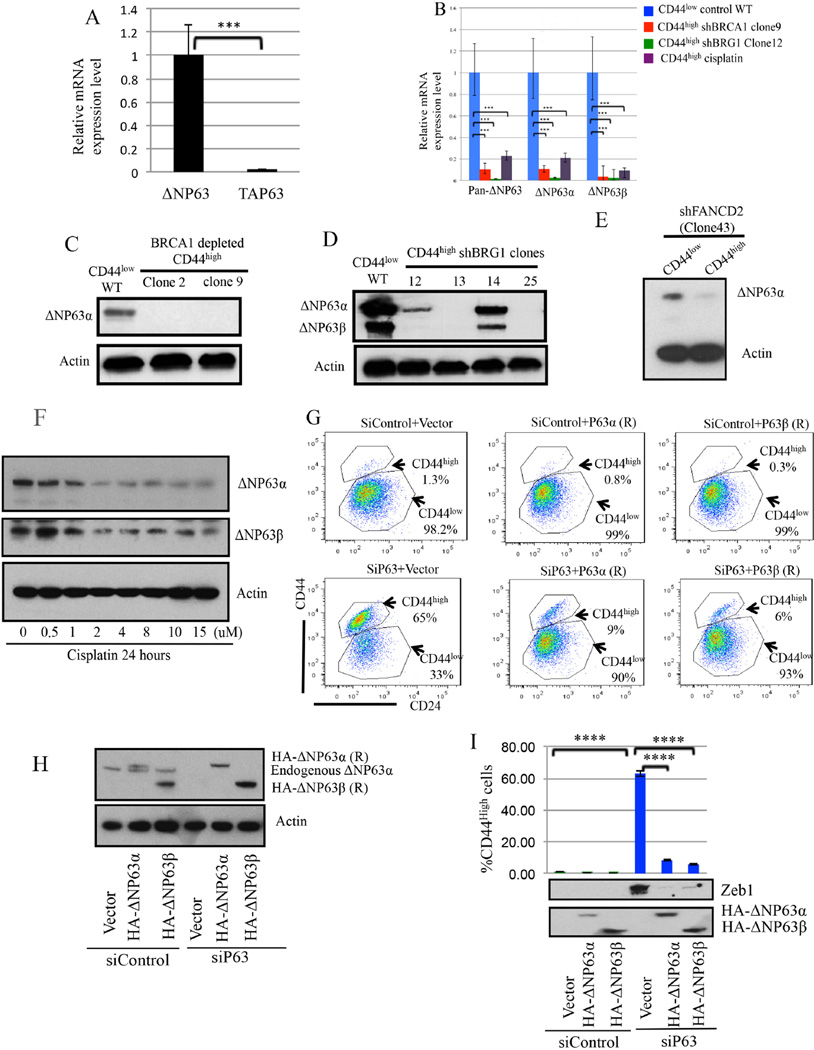
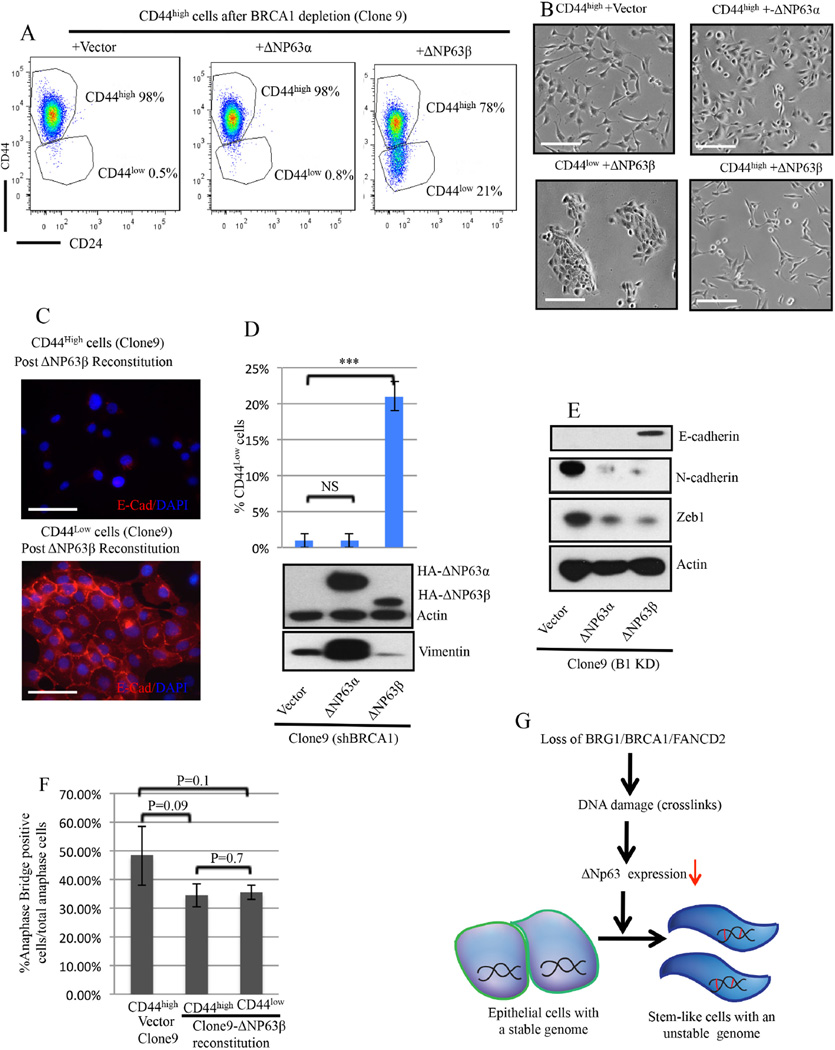
An abnormal differentiation state is common in BRCA1-deficient mammary epithelial cells, but the underlying mechanism is unclear. Here, we report a convergence between DNA repair and normal, cultured human mammary epithelial (HME) cell differentiation. Surprisingly, depleting BRCA1 or FANCD2 (Fanconi anemia [FA] proteins) or BRG1, a mSWI/SNF subunit, caused HME cells to undergo spontaneous epithelial-to-mesenchymal transition (EMT) and aberrant differentiation. This also occurred when wild-type HMEs were exposed to chemicals that generate DNA interstrand crosslinks (repaired by FA proteins), but not in response to double-strand breaks. Suppressed expression of ΔNP63 also occurred in each of these settings, an effect that links DNA damage to the aberrant differentiation outcome. Taken together with somatic breast cancer genome data, these results point to a breakdown in a BRCA/FA-mSWI/SNF-ΔNP63-mediated DNA repair and differentiation maintenance process in mammary epithelial cells that may contribute to sporadic breast cancer development.
Keywords: BRCA1; BRG1; EMT; FANCD2; breast cancer; cisplatin; crosslink repair.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Adorno M, Cordenonsi M, Montagner M, Dupont S, Wong C, Hann B, Solari A, Bobisse S, Rondina MB, Guzzardo V, et al. A Mutant-p53/Smad complex opposes p63 to empower TGFbeta-induced metastasis. Cell. 2009;137:87–98. - PubMed
-
- Aguilera A, Garcia-Muse T. R loops: from transcription byproducts to threats to genome stability. Molecular cell. 2012;46:115–124. - PubMed
-
- Bochar DA, Wang L, Beniya H, Kinev A, Xue Y, Lane WS, Wang W, Kashanchi F, Shiekhattar R. BRCA1 is associated with a human SWI/SNF-related complex: linking chromatin remodeling to breast cancer. Cell. 2000;102:257–265. - PubMed
-
- Buckley NE, Conlon SJ, Jirstrom K, Kay EW, Crawford NT, O’Grady A, Sheehan K, Mc Dade SS, Wang CW, McCance DJ, et al. The DeltaNp63 proteins are key allies of BRCA1 in the prevention of basal-like breast cancer. Cancer research. 2011;71:1933–1944. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
