

Competitive antagonists facilitate the recovery from desensitization of α1β2γ2 GABAA receptors expressed in Xenopus oocytes
- PMID: 27374488
- PMCID: PMC4973384
- DOI: 10.1038/aps.2016.50
Competitive antagonists facilitate the recovery from desensitization of α1β2γ2 GABAA receptors expressed in Xenopus oocytes
Abstract
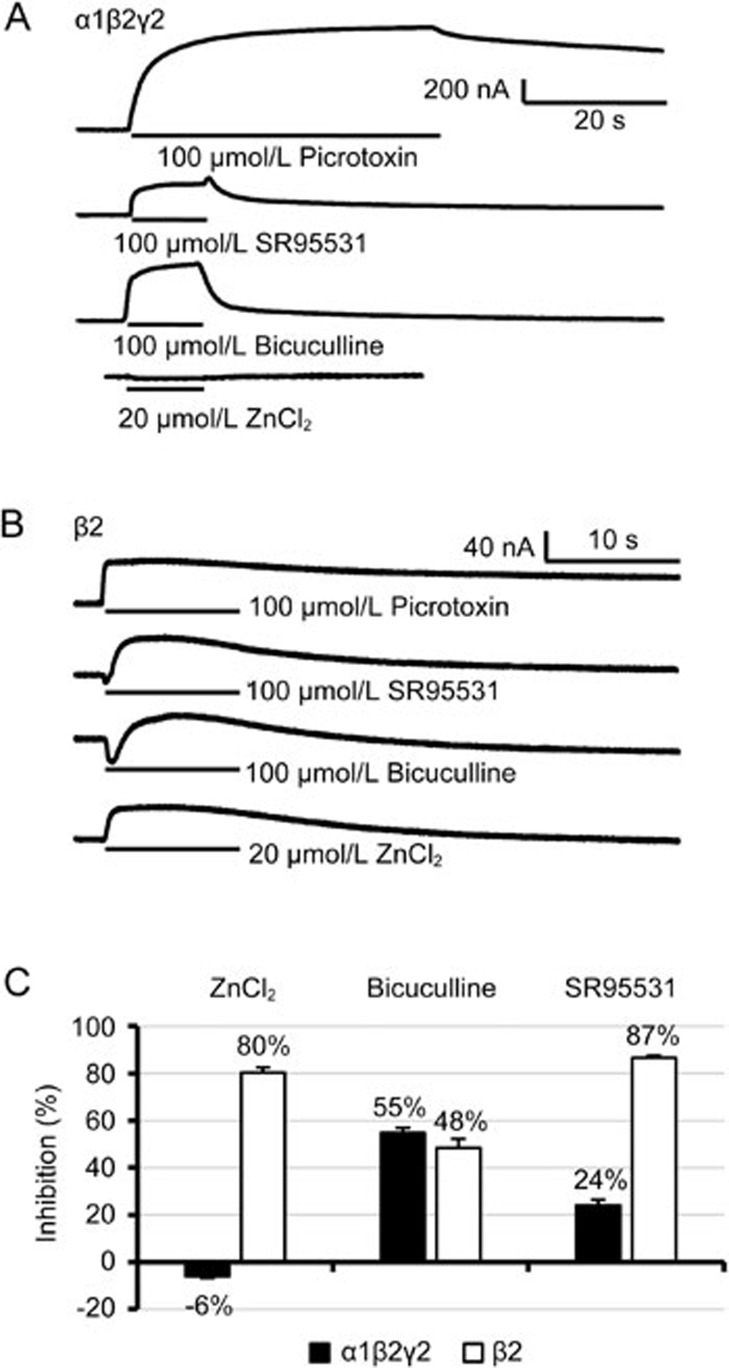
Aim: The continuous presence of an agonist drives its receptor into a refractory state, termed desensitization. In this study, we tested the hypothesis that a competitive antagonist, SR95531, could facilitate the recovery of α1β2γ2 GABAA receptor from functional desensitization.
Methods: α1β2γ2 GABAA receptors were expressed in Xenopus oocytes. GABA-evoked currents were recorded using two-electrode voltage-clamp technique. Drugs were applied through perfusion.
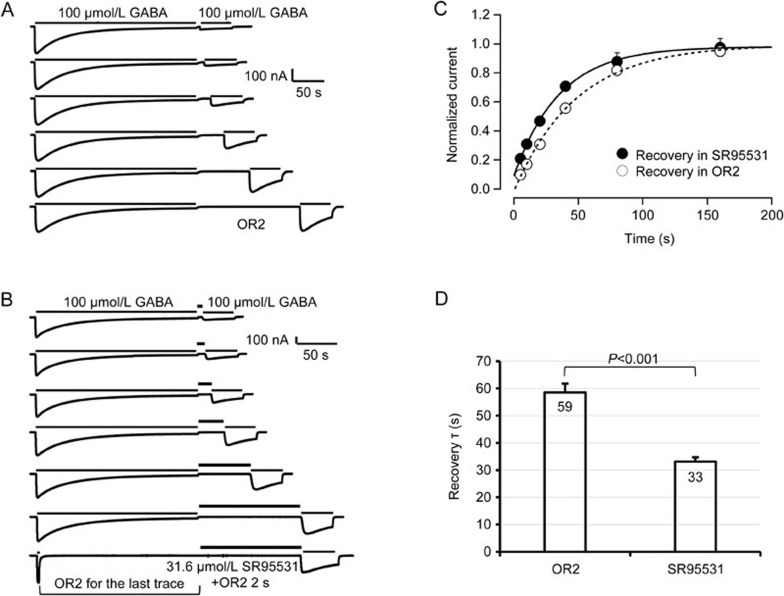
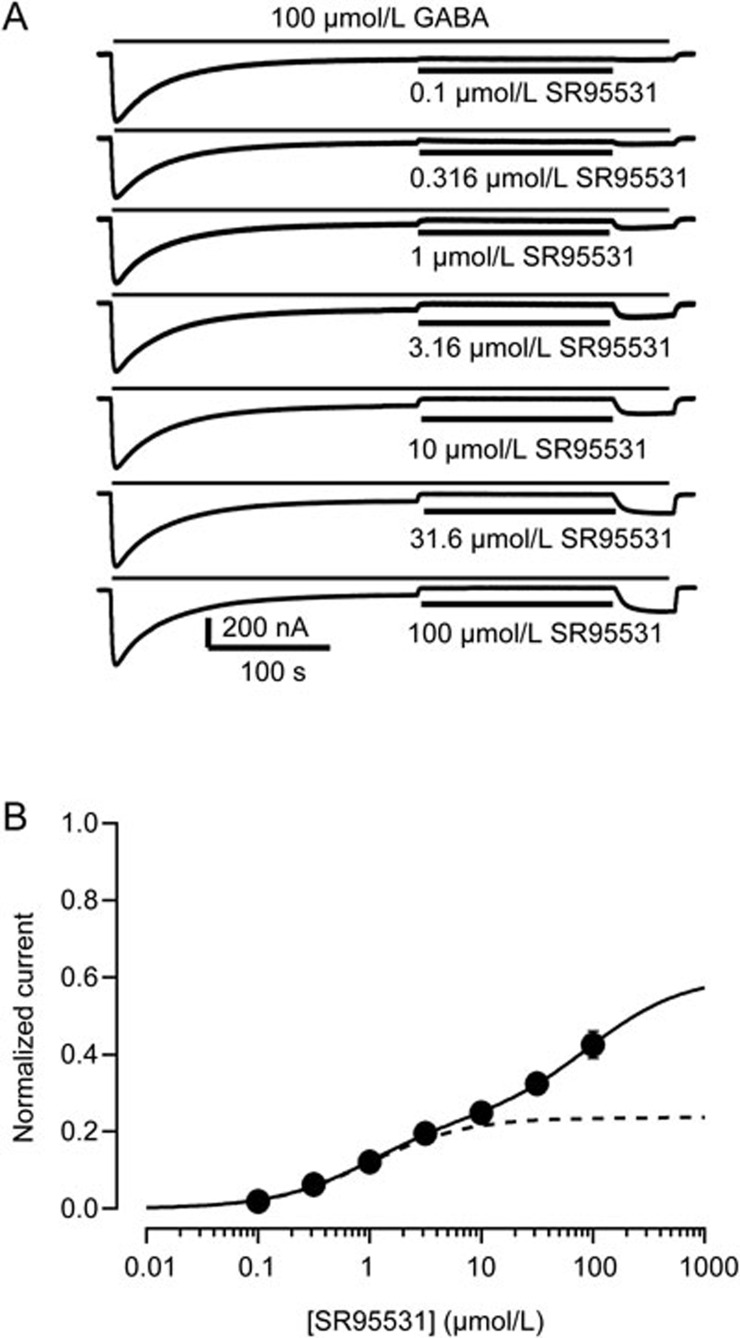
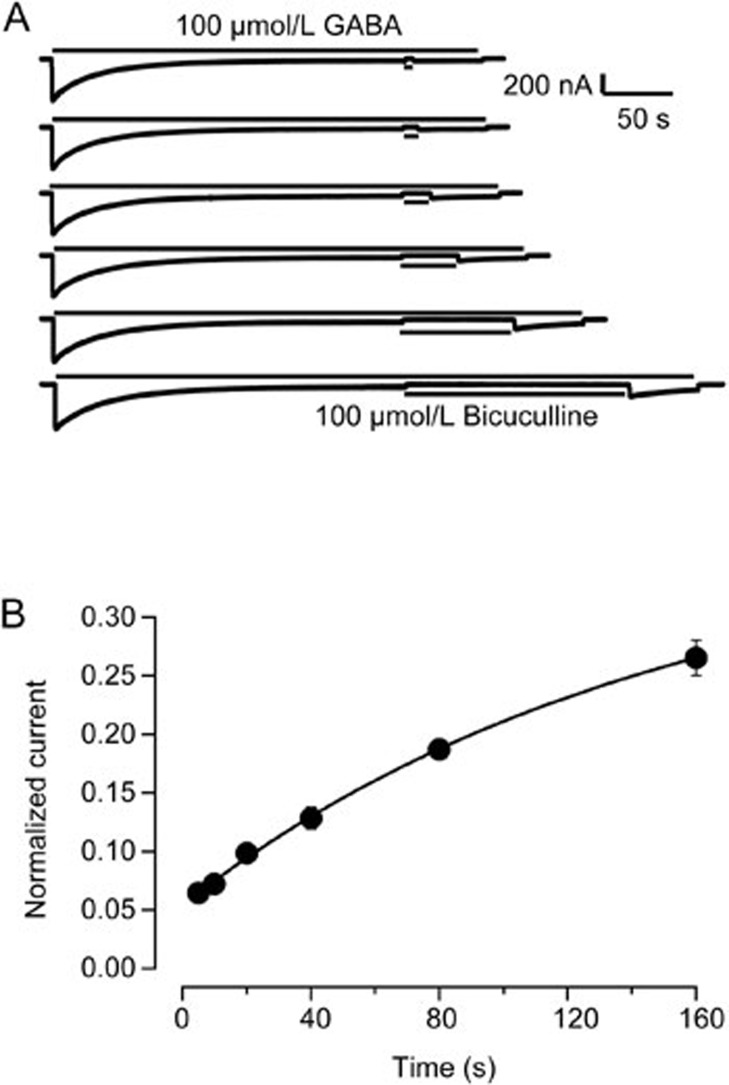
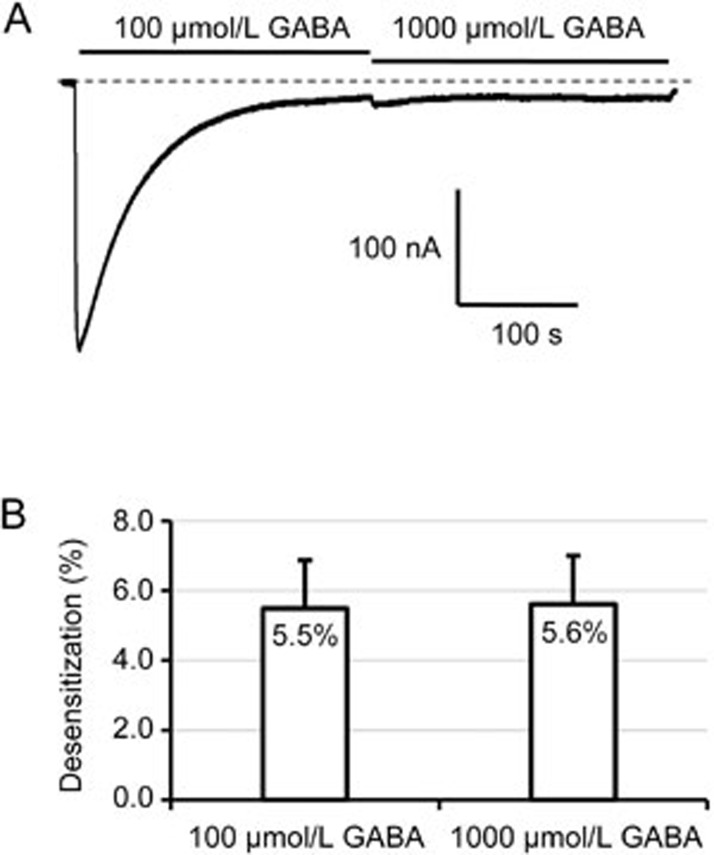
Results: Long application of GABA (100 μmol/L) evoked a large peak current followed by a small amplitude steady-state current (desensitization). Co-application of SR95531 during the desensitization caused a larger rebound of GABA current after removal of SR95531. Furthermore, application of SR95531 after removal of GABA increased the rate of receptor recovery from desensitization, and the recovery time constant was decreased from 59±3.2 s to 33±1.6 s. SR95531-facilitated receptor recovery from desensitization was dependent on the perfusion duration of SR95531. It was also dependent on the concentration of SR95531, and the curve fitting with Hill equation revealed two potency components, which were similar to the two potency components in inhibition of the steady-state current by SR95531. Bicuculline caused similar facilitation of desensitization recovery.
Conclusion: SR95531 facilitates α1β2γ2 GABAA receptor recovery from desensitization, possibly through two mechanisms: binding to the desensitized receptor and converting it to the non-desensitized state, and binding to the resting state receptor and preventing re-desensitization.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
