

The draft genome of MD-2 pineapple using hybrid error correction of long reads
- PMID: 27374615
- PMCID: PMC5066169
- DOI: 10.1093/dnares/dsw026
The draft genome of MD-2 pineapple using hybrid error correction of long reads
Abstract
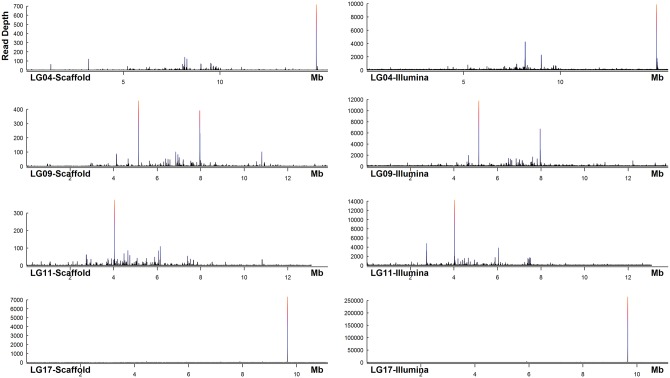
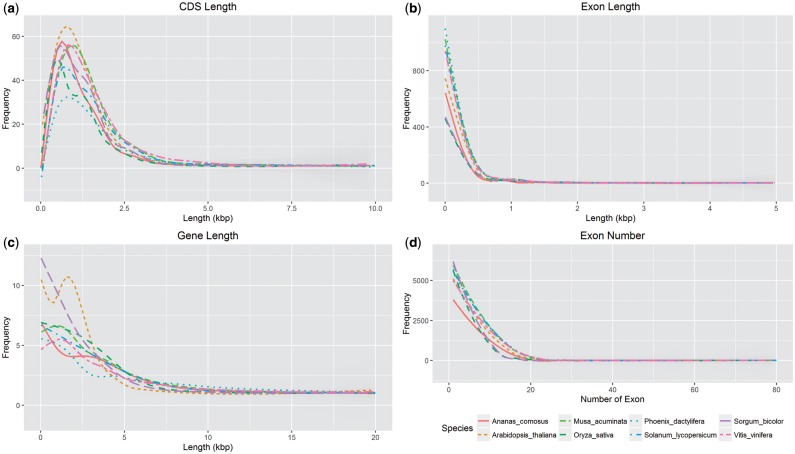
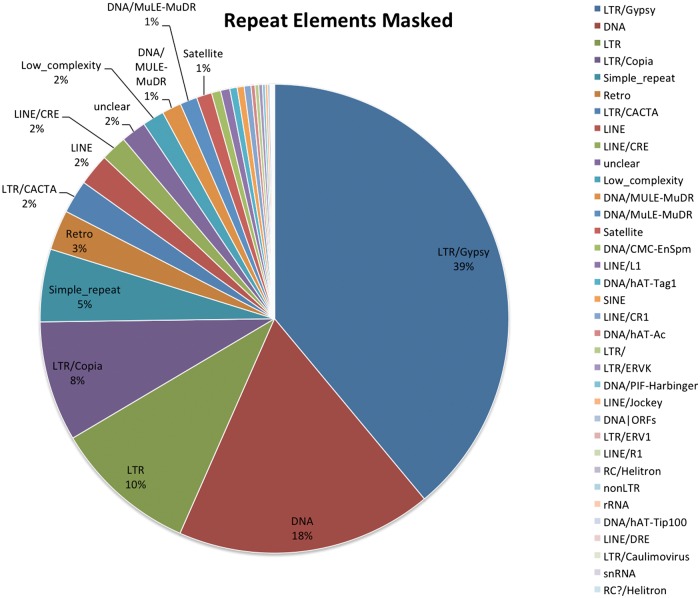
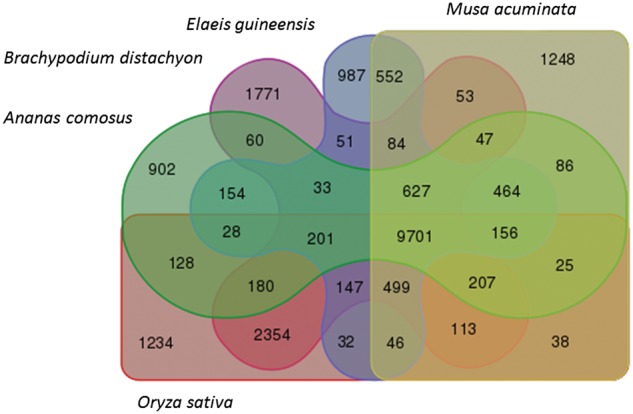
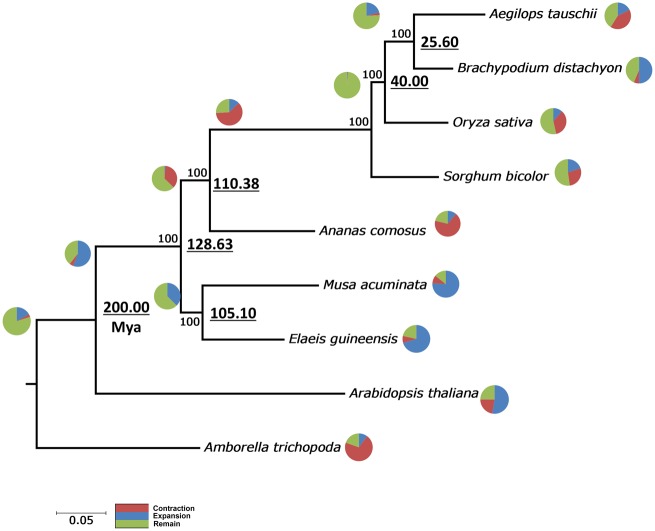
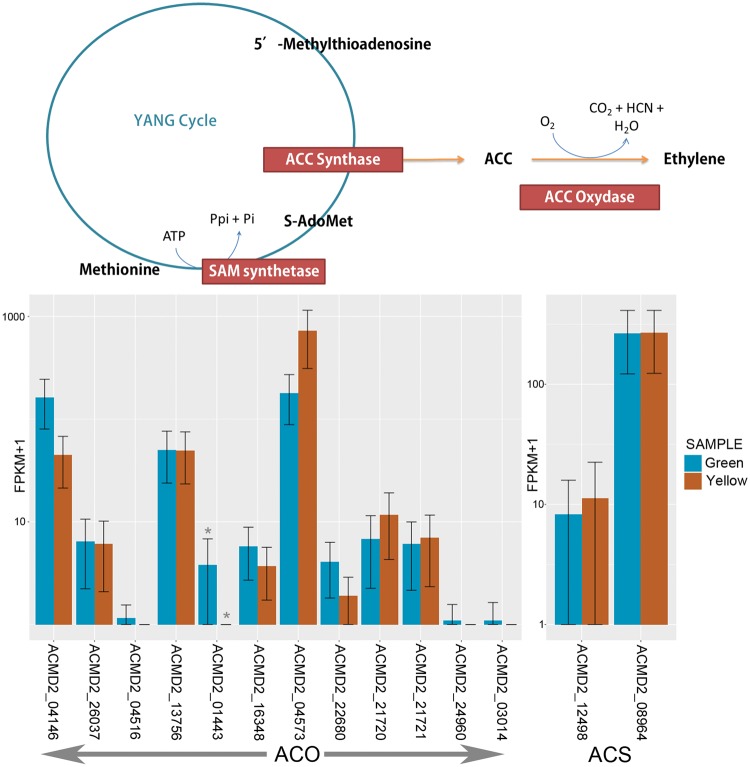
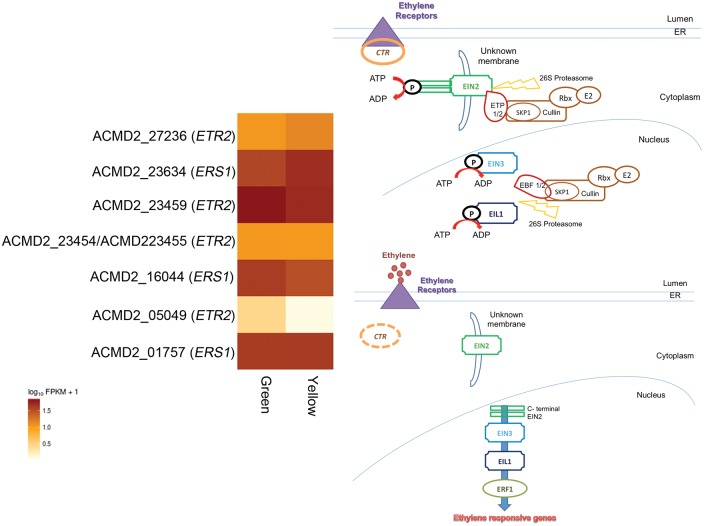
The introduction of the elite pineapple variety, MD-2, has caused a significant market shift in the pineapple industry. Better productivity, overall increased in fruit quality and taste, resilience to chilled storage and resistance to internal browning are among the key advantages of the MD-2 as compared with its previous predecessor, the Smooth Cayenne. Here, we present the genome sequence of the MD-2 pineapple (Ananas comosus (L.) Merr.) by using the hybrid sequencing technology from two highly reputable platforms, i.e. the PacBio long sequencing reads and the accurate Illumina short reads. Our draft genome achieved 99.6% genome coverage with 27,017 predicted protein-coding genes while 45.21% of the genome was identified as repetitive elements. Furthermore, differential expression of ripening RNASeq library of pineapple fruits revealed ethylene-related transcripts, believed to be involved in regulating the process of non-climacteric pineapple fruit ripening. The MD-2 pineapple draft genome serves as an example of how a complex heterozygous genome is amenable to whole genome sequencing by using a hybrid technology that is both economical and accurate. The genome will make genomic applications more feasible as a medium to understand complex biological processes specific to pineapple.
Keywords: fruit ripening; hybrid assembly; pineapple; plant genome.
© The Author 2016. Published by Oxford University Press on behalf of Kazusa DNA Research Institute.
Figures
References
-
- Arumuganathan K., Earle E.D. 1991, Nuclear DNA content of some important plant species, Plant Mol. Biol. Report., 9, 415.
-
- Moore S. 2002, Use of genomics tools to isolate key ripening genes and analyse fruit maturation in tomato, J. Exp. Bot., 53, 2023–30. - PubMed
-
- Cherian S., Figueroa C.R., Nair H. 2014, ‘ Movers and shakers’ in the regulation of fruit ripening: a cross-dissection of climacteric versus non-climacteric fruit, J. Exp. Bot., 65, 4705–22. - PubMed
-
- Golding J.B., Shearer D., Wyllie S.G., McGlasson W. 1998, Application of 1-MCP and propylene to identify ethylene-dependent ripening processes in mature banana fruit, Postharvest Biol. Technol., 14, 87–98.
LinkOut - more resources
Full Text Sources
Other Literature Sources
