

Hierarchy within the mammary STAT5-driven Wap super-enhancer
- PMID: 27376239
- PMCID: PMC4963296
- DOI: 10.1038/ng.3606
Hierarchy within the mammary STAT5-driven Wap super-enhancer
Abstract
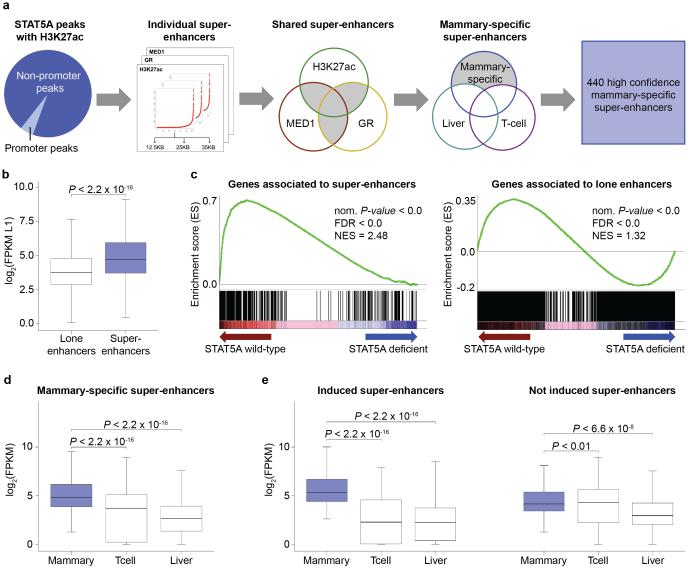
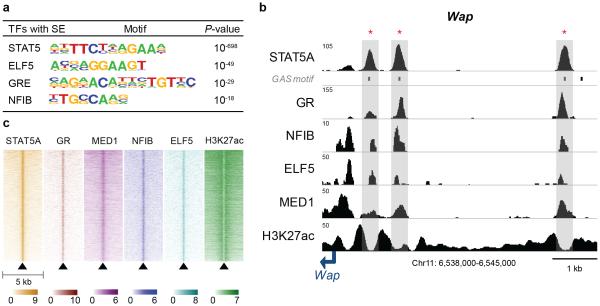
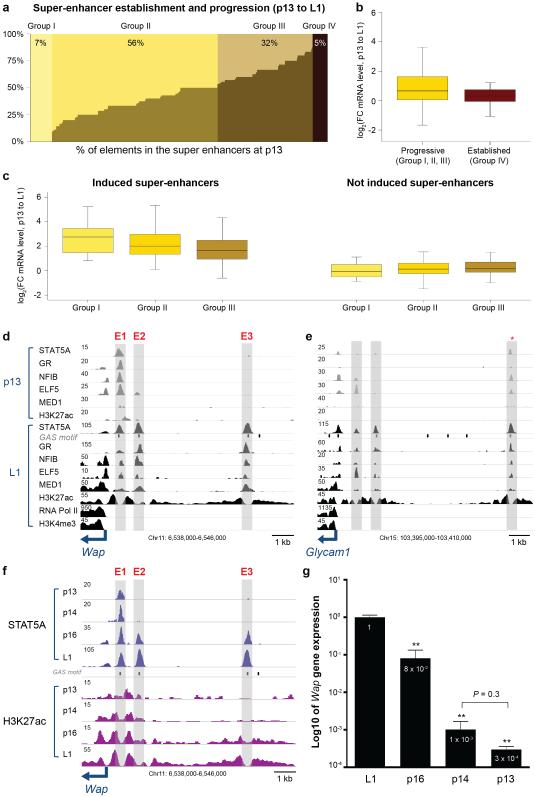
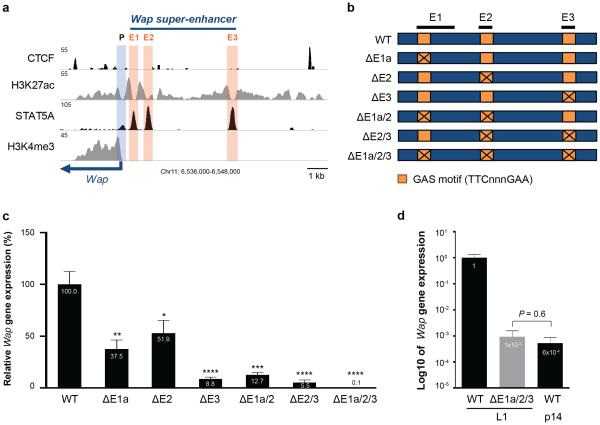
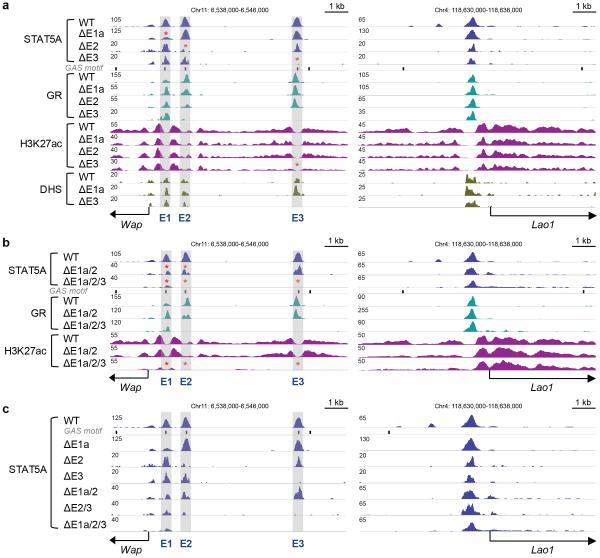
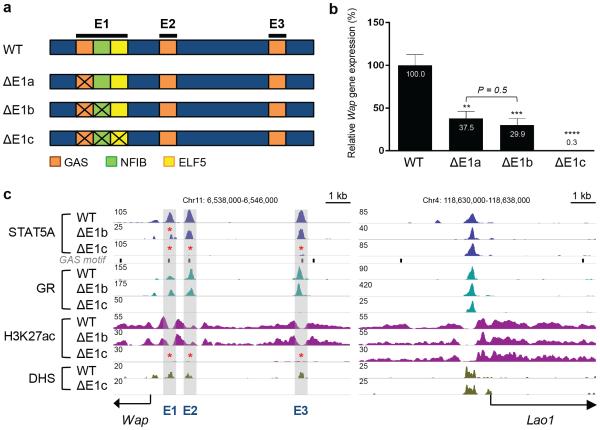
Super-enhancers comprise dense transcription factor platforms highly enriched for active chromatin marks. A paucity of functional data led us to investigate the role of super-enhancers in the mammary gland, an organ characterized by exceptional gene regulatory dynamics during pregnancy. ChIP-seq analysis for the master regulator STAT5A, the glucocorticoid receptor, H3K27ac and MED1 identified 440 mammary-specific super-enhancers, half of which were associated with genes activated during pregnancy. We interrogated the Wap super-enhancer, generating mice carrying mutations in STAT5-binding sites within its constituent enhancers. Individually, the most distal site displayed the greatest enhancer activity. However, combinatorial mutation analysis showed that the 1,000-fold induction in gene expression during pregnancy relied on all enhancers. Disabling the binding sites of STAT5, NFIB and ELF5 in the proximal enhancer incapacitated the entire super-enhancer. Altogether, these data suggest a temporal and functional enhancer hierarchy. The identification of mammary-specific super-enhancers and the mechanistic exploration of the Wap locus provide insights into the regulation of cell-type-specific expression of hormone-sensing genes.
Figures
Comment in
-
Is a super-enhancer greater than the sum of its parts?Nat Genet. 2016 Dec 28;49(1):2-3. doi: 10.1038/ng.3759. Nat Genet. 2016. PMID: 28029159 Free PMC article. No abstract available.
References
-
- Hennighausen L, Robinson GW. Information networks in the mammary gland. Nat Rev Mol Cell Biol. 2005;6:715–25. - PubMed
-
- Lydon JP, et al. Mice lacking progesterone receptor exhibit pleiotropic reproductive abnormalities. Genes Dev. 1995;9:2266–78. - PubMed
-
- Ormandy CJ, et al. Null mutation of the prolactin receptor gene produces multiple reproductive defects in the mouse. Genes Dev. 1997;11:167–78. - PubMed
-
- Waller A, Adunski A, Hershkowitz M. Terminal dehydration and intravenous fluids. Lancet. 1991;337:745. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
