

Structural studies of N-terminal mutants of Connexin 26 and Connexin 32 using (1)H NMR spectroscopy
- PMID: 27378082
- PMCID: PMC5051353
- DOI: 10.1016/j.abb.2016.06.019
Structural studies of N-terminal mutants of Connexin 26 and Connexin 32 using (1)H NMR spectroscopy
Abstract
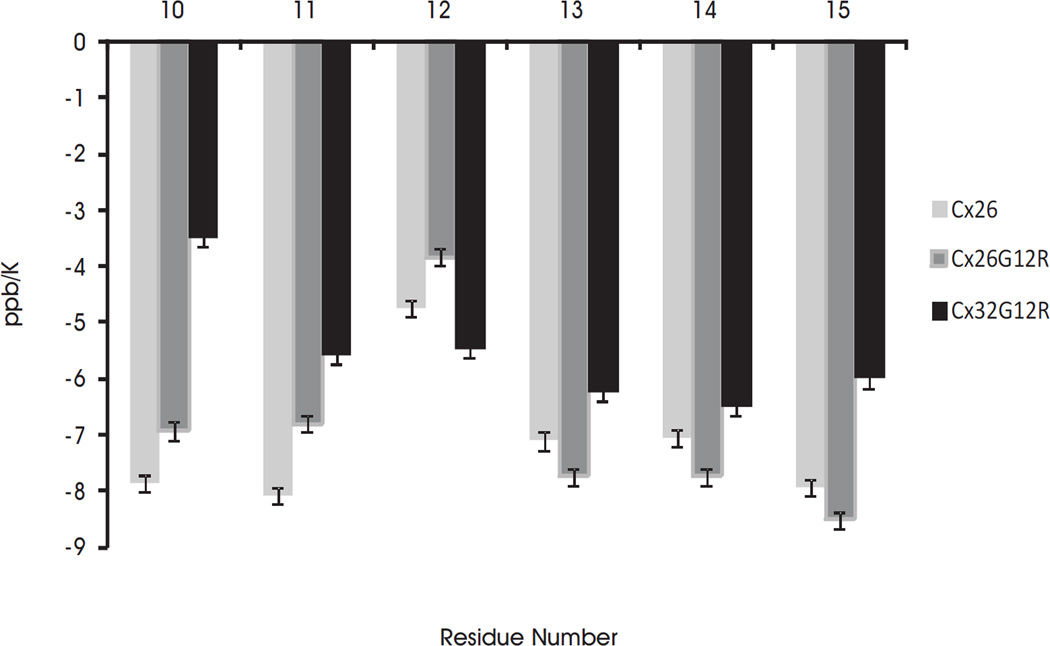
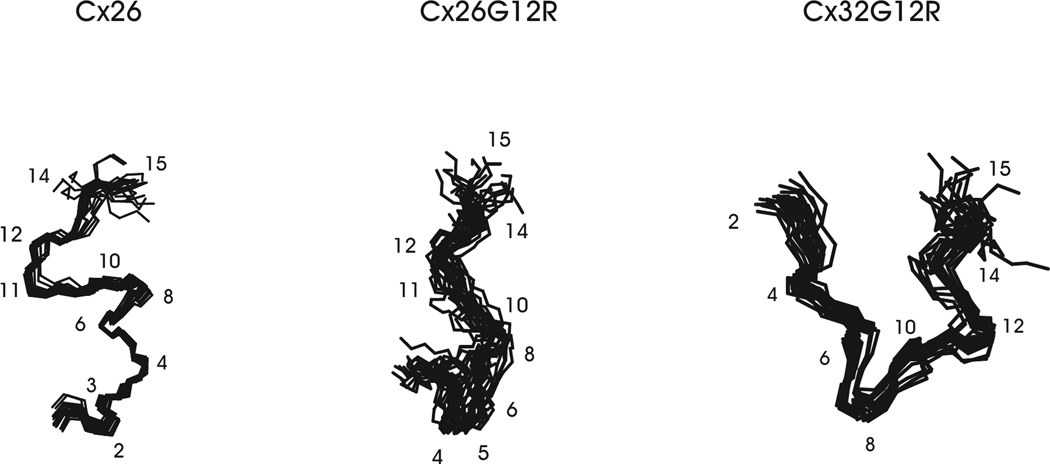
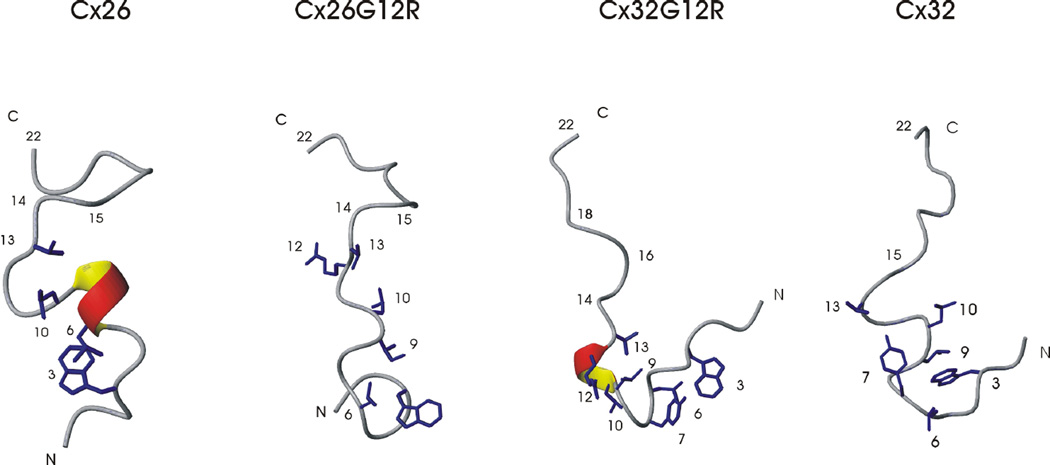
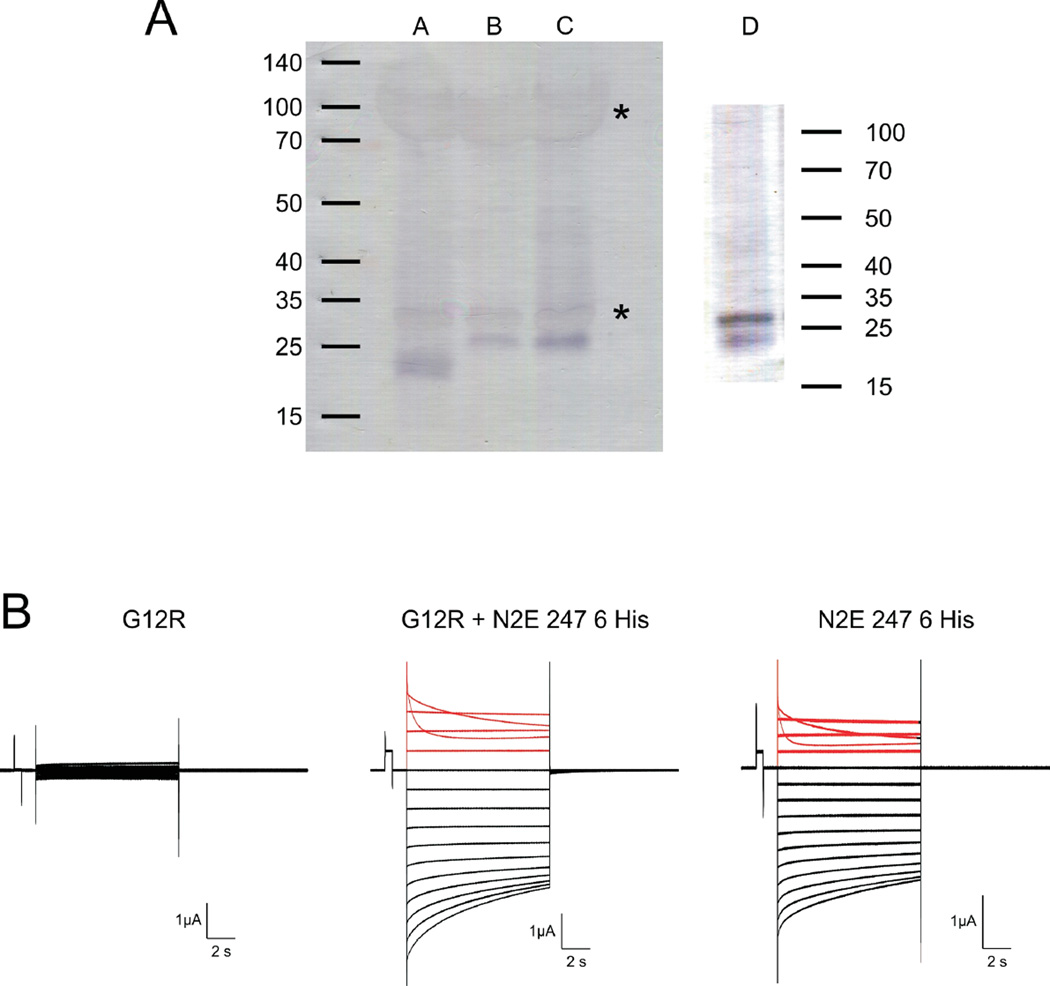
Alterations in gap junctions underlie the etiologies of syndromic deafness (KID) and Charcot-Marie Tooth disease (CMTX). Functional gap junctions are composed of connexin molecules with N-termini containing a flexible turn around G12, inserting the N-termini into the channel pore allowing voltage gating. The loss of this turn correlates with loss of Connexin 32 (Cx32) function by impaired trafficking to the cell membrane. Using (1)H NMR we show the N-terminus of a syndromic deafness mutation Cx26G12R, producing "leaky channels", contains a turn around G12 which is less structured and more flexible than wild-type. In contrast, the N-terminal structure of the same mutation in Cx32 chimera, Cx32*43E1G12R shows a larger constricted turn and no membrane current expression but forms membrane inserted hemichannels. Their function was rescued by formation of heteromeric channels with wild type subunits. We suggest the inflexible Cx32G12R N-terminus blocks ion conduction in homomeric channels and this channel block is relieved by incorporation of wild type subunits. In contrast, the increased open probability of Cx26G12R hemichannels is likely due to the addition of positive charge in the channel pore changing pore electrostatics and impairing hemichannel regulation by Ca(2+). These results provide mechanistic information on aberrant channel activity observed in disease.
Keywords: Connexin; Deafness; NMR; Peptides; Structure-function.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Structure of the amino terminus of a gap junction protein.Arch Biochem Biophys. 2000 Sep 15;381(2):181-90. doi: 10.1006/abbi.2000.1989. Arch Biochem Biophys. 2000. PMID: 11032405
-
Stoichiometry of transjunctional voltage-gating polarity reversal by a negative charge substitution in the amino terminus of a connexin32 chimera.J Gen Physiol. 2000 Jul 1;116(1):13-31. doi: 10.1085/jgp.116.1.13. J Gen Physiol. 2000. PMID: 10871637 Free PMC article.
-
Structural studies of N-terminal mutants of connexin 32 using (1)H NMR spectroscopy.Arch Biochem Biophys. 2012 Oct 1;526(1):1-8. doi: 10.1016/j.abb.2012.05.027. Epub 2012 Jun 14. Arch Biochem Biophys. 2012. PMID: 22705201 Free PMC article.
-
Mutations in connexin 32: the molecular and biophysical bases for the X-linked form of Charcot-Marie-Tooth disease.Brain Res Brain Res Rev. 2000 Apr;32(1):203-14. doi: 10.1016/s0165-0173(99)00082-x. Brain Res Brain Res Rev. 2000. PMID: 10751671 Review.
Cited by
-
Simulation of gap junction formation reveals critical role of Cys disulfide redox state in connexin hemichannel docking.Cell Commun Signal. 2024 Mar 18;22(1):185. doi: 10.1186/s12964-023-01439-z. Cell Commun Signal. 2024. PMID: 38500186 Free PMC article.
-
Role of ROS/RNS in Preeclampsia: Are Connexins the Missing Piece?Int J Mol Sci. 2020 Jun 30;21(13):4698. doi: 10.3390/ijms21134698. Int J Mol Sci. 2020. PMID: 32630161 Free PMC article. Review.
-
Connexin Gap Junction Channels and Hemichannels: Insights from High-Resolution Structures.Biology (Basel). 2024 Apr 26;13(5):298. doi: 10.3390/biology13050298. Biology (Basel). 2024. PMID: 38785780 Free PMC article. Review.
-
Connexons Coupling to Gap Junction Channel: Potential Role for Extracellular Protein Stabilization Centers.Biomolecules. 2021 Dec 30;12(1):49. doi: 10.3390/biom12010049. Biomolecules. 2021. PMID: 35053197 Free PMC article.
-
Functional Consequences of Pathogenic Variants of the GJB2 Gene (Cx26) Localized in Different Cx26 Domains.Biomolecules. 2023 Oct 13;13(10):1521. doi: 10.3390/biom13101521. Biomolecules. 2023. PMID: 37892203 Free PMC article. Review.
References
-
- Maeda S, Nakagawa S, Suga M, Yamashita E, Oshima A, Fujiyoshi Y, Tsukihara T. Structure of the connexin 26 gap junction channel at 3.5Å resolution. Nature. 2011;458:597–602. - PubMed
-
- Bennett MVL, Zukin RS. Electrical Coupling and Neuronal Synchronization in the Mammalian Brain. Neuron. 2004;41:495–511. - PubMed
-
- Potolicchio I, Cigliola V, Velazquez-Garcia S, Klee P, Valjevac A, Kapic D, Cosovic E, Lepara O, Hadzovic-Dzuvo A, Mornjacovic Z, Meda P. Connexin-dependent signaling in neuro-hormonal systems. (BBA) - Biomembranes. 2012;1818:1919–1936. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
