

Identification of plexin A4 as a novel clusterin receptor links two Alzheimer's disease risk genes
- PMID: 27378688
- PMCID: PMC5179943
- DOI: 10.1093/hmg/ddw188
Identification of plexin A4 as a novel clusterin receptor links two Alzheimer's disease risk genes
Abstract
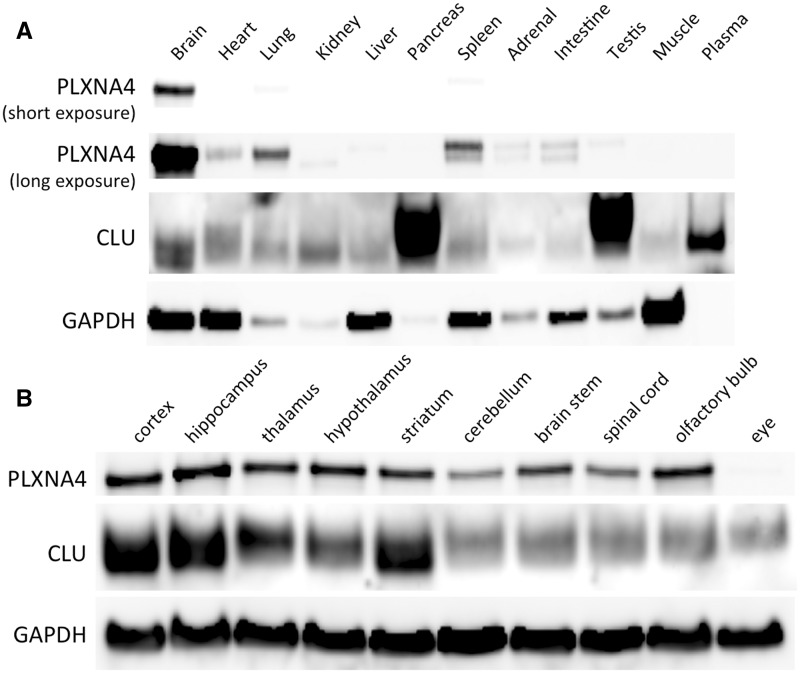
Although abundant genetic and biochemical evidence strongly links Clusterin (CLU) to Alzheimer disease (AD) pathogenesis, the receptor for CLU within the adult brain is currently unknown. Using unbiased approaches, we identified Plexin A4 (PLXNA4) as a novel, high-affinity receptor for CLU in the adult brain. PLXNA4 protein expression was high in brain with much lower levels in peripheral organs. CLU protein levels were significantly elevated in the cerebrospinal fluid (CSF) of Plxna4-/- mice and, in humans, CSF levels of CLU were also associated with PLXNA4 genotype. Human AD brains had significantly increased the levels of CLU protein but decreased levels of PLXNA4 by ∼50%. To determine whether PLXNA4 levels influenced cognition, we analyzed the behaviour of Plxna4+/+, Plxna4+/-, and Plxna4-/- mice. In comparison to WT controls, both Plxna4+/- and Plxna4-/- mice were hyperactive in the open field assay while Plxna4-/- mice displayed a hyper-exploratory (low-anxiety phenotype) in the elevated plus maze. Importantly, both Plxna4+/- and Plxna4-/- mice displayed prominent deficits in learning and memory in the contextual fear-conditioning paradigm. Thus, even a 50% reduction in the level of PLXNA4 is sufficient to cause memory impairments, raising the possibility that memory problems seen in AD patients could be due to reductions in the level of PLXNA4. Both CLU and PLXNA4 have been genetically associated with AD risk and our data thus provide a direct relationship between two AD risk genes. Our data suggest that increasing the levels of PLXNA4 or targeting CLU-PLXNA4 interactions may have therapeutic value in AD.
© The Author 2016. Published by Oxford University Press.
Figures






References
-
- DeMattos R.B., Cirrito J.R., Parsadanian M., May P.C., O’Dell M.A., Taylor J.W., Harmony J.A.K., Aronow B.J., Bales K.R., Paul S.M., et al. (2004) ApoE and clusterin cooperatively suppress Abeta levels and deposition: evidence that ApoE regulates extracellular Abeta metabolism in vivo. Neuron, 41, 193–202. - PubMed
-
- DeMattos R.B., O’dell M.A., Parsadanian M., Taylor J.W., Harmony J.A.K., Bales K.R., Paul S.M., Aronow B.J., Holtzman D.M. (2002) Clusterin promotes amyloid plaque formation and is critical for neuritic toxicity in a mouse model of Alzheimer’s disease. Proc. Natl. Acad. Sci. U. S. A, 99, 10843–10848. - PMC - PubMed
-
- Lambert J.C., Heath S., Even G., Campion D., Sleegers K., Hiltunen M., Combarros O., Zelenika D., Bullido M.J., Tavernier B., et al. (2009) Genome-wide association study identifies variants at CLU and CR1 associated with Alzheimer’s disease. Nat. Genet., 41, 1094–1099. - PubMed
-
- Gajera C.R., Emich H., Lioubinski O., Christ A., Beckervordersandforth-Bonk R., Yoshikawa K., Bachmann S., Christensen E.I., Götz M., Kempermann G., et al. (2010) LRP2 in ependymal cells regulates BMP signaling in the adult neurogenic niche. J. Cell Sci., 123, 1922–1930. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
