

Mechanistic insights into selective autophagy pathways: lessons from yeast
- PMID: 27381245
- PMCID: PMC5549613
- DOI: 10.1038/nrm.2016.74
Mechanistic insights into selective autophagy pathways: lessons from yeast
Abstract
Autophagy has burgeoned rapidly as a field of study because of its evolutionary conservation, the diversity of intracellular cargoes degraded and recycled by this machinery, the mechanisms involved, as well as its physiological relevance to human health and disease. This self-eating process was initially viewed as a non-selective mechanism used by eukaryotic cells to degrade and recycle macromolecules in response to stress; we now know that various cellular constituents, as well as pathogens, can also undergo selective autophagy. In contrast to non-selective autophagy, selective autophagy pathways rely on a plethora of selective autophagy receptors (SARs) that recognize and direct intracellular protein aggregates, organelles and pathogens for specific degradation. Although SARs themselves are not highly conserved, their modes of action and the signalling cascades that activate and regulate them are. Recent yeast studies have provided novel mechanistic insights into selective autophagy pathways, revealing principles of how various cargoes can be marked and targeted for selective degradation.
Conflict of interest statement
The authors declare no competing interests.
Figures
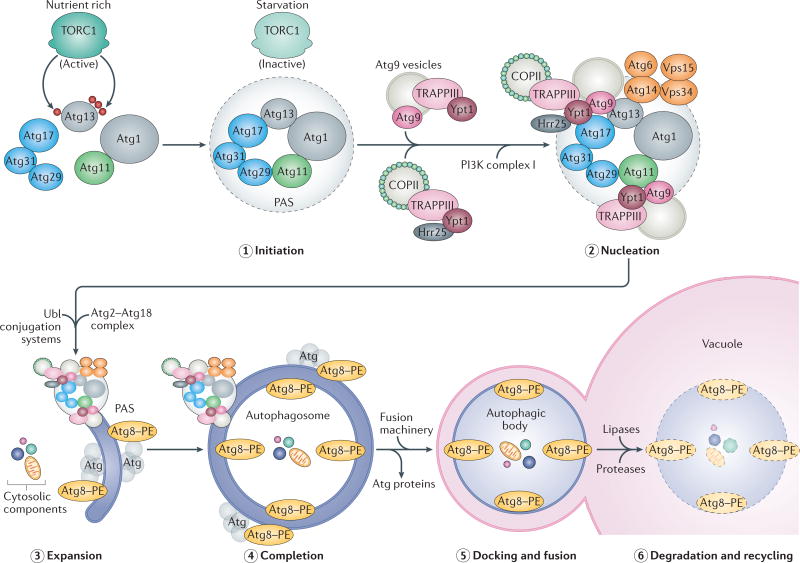
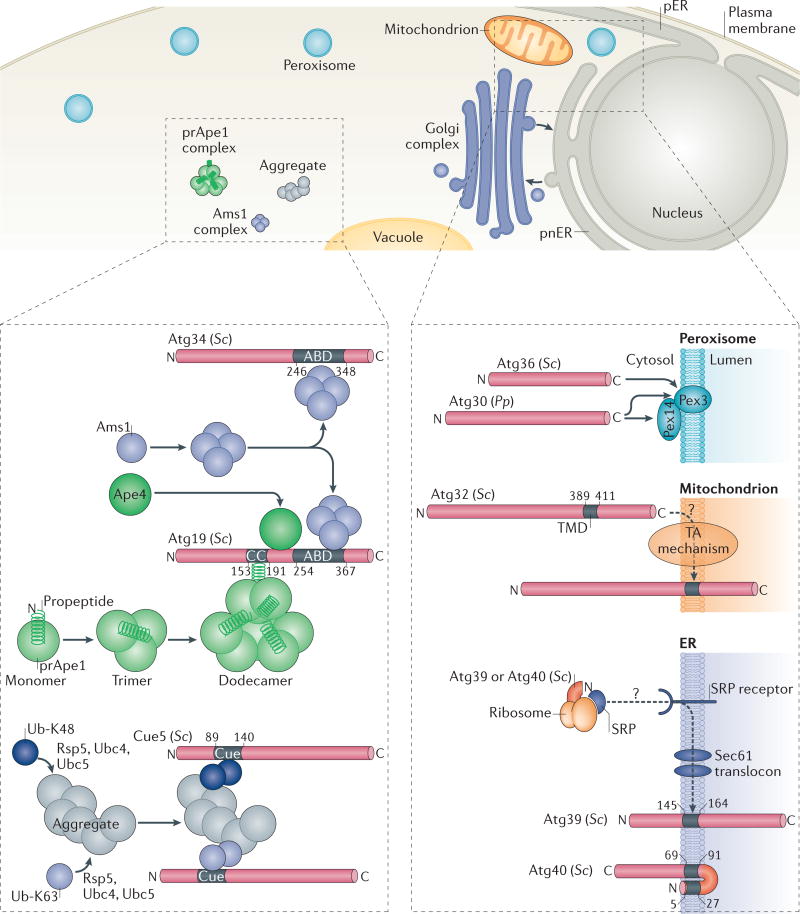
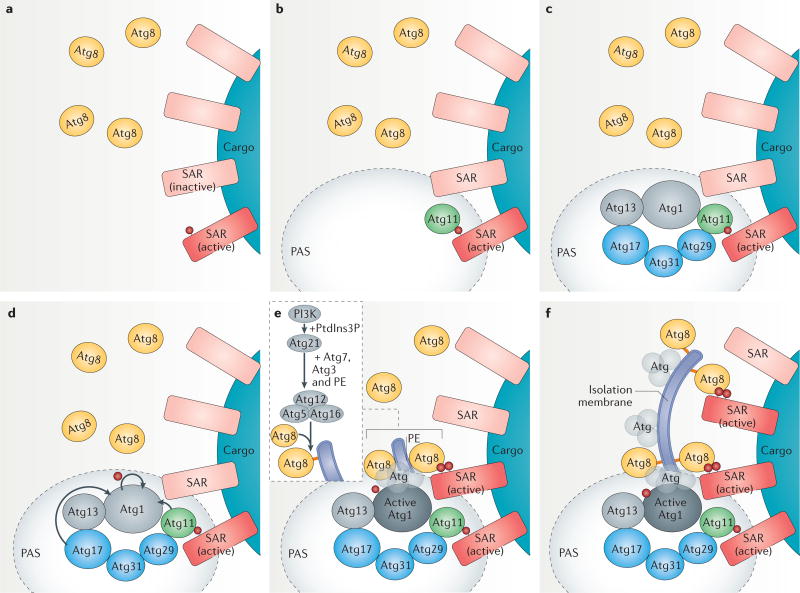
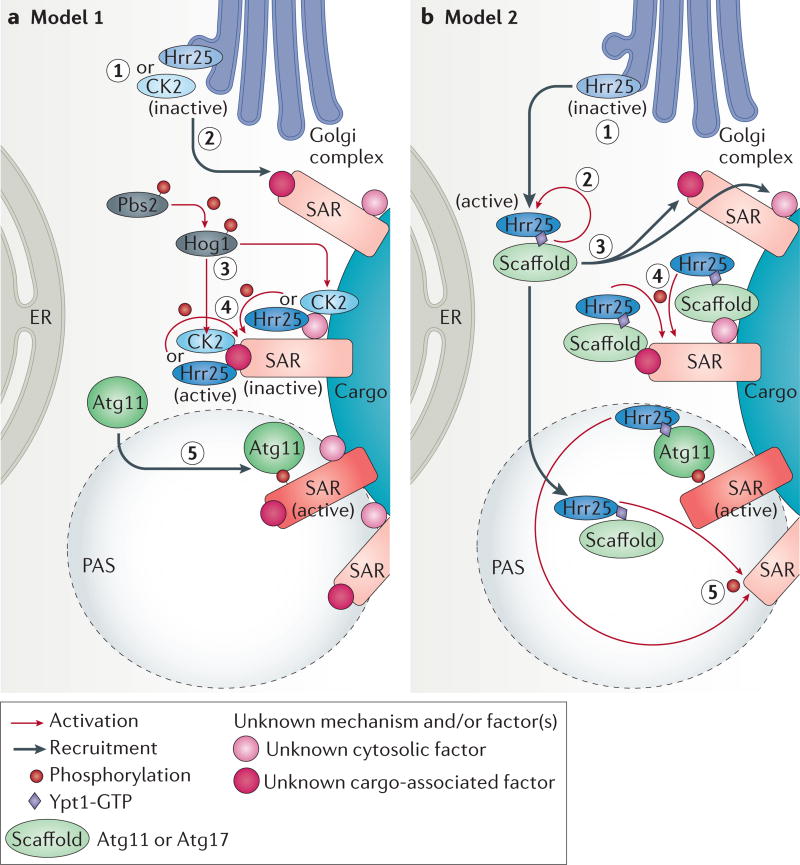
References
-
- Choi AM, Ryter SW, Levine B. Autophagy in human health and disease. N. Engl. J. Med. 2013;368:651–662. - PubMed
-
- Gozuacik D, Kimchi A. Autophagy as a cell death and tumor suppressor mechanism. Oncogene. 2004;23:2891–2906. - PubMed
-
- Rubinsztein DC, Marino G, Kroemer G. Autophagy and aging. Cell. 2011;146:682–695. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
