

Infectivity of Plasmodium falciparum in Malaria-Naive Individuals Is Related to Knob Expression and Cytoadherence of the Parasite
- PMID: 27382019
- PMCID: PMC4995910
- DOI: 10.1128/IAI.00414-16
Infectivity of Plasmodium falciparum in Malaria-Naive Individuals Is Related to Knob Expression and Cytoadherence of the Parasite
Abstract
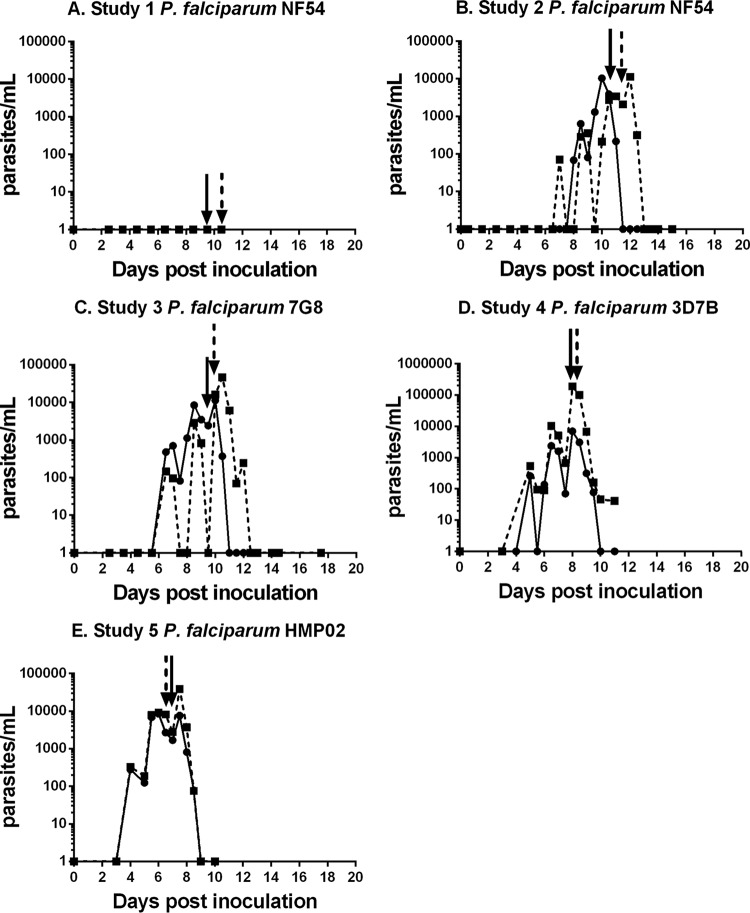
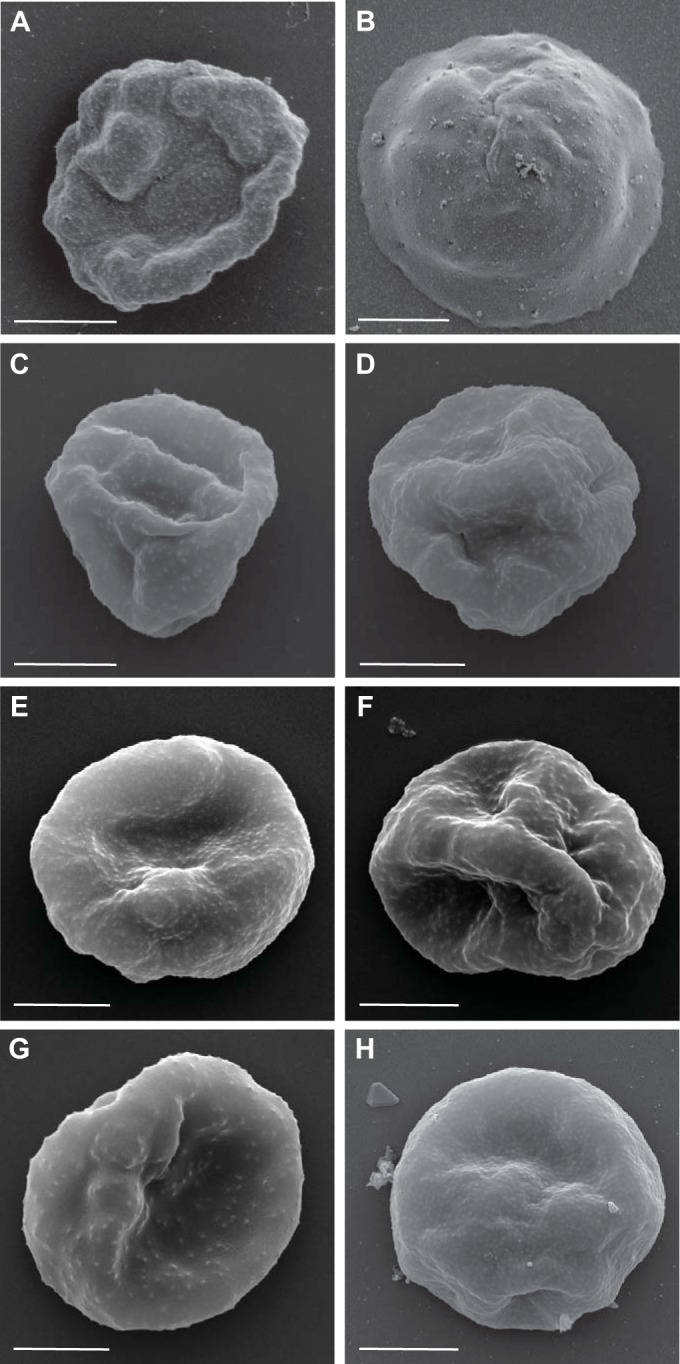
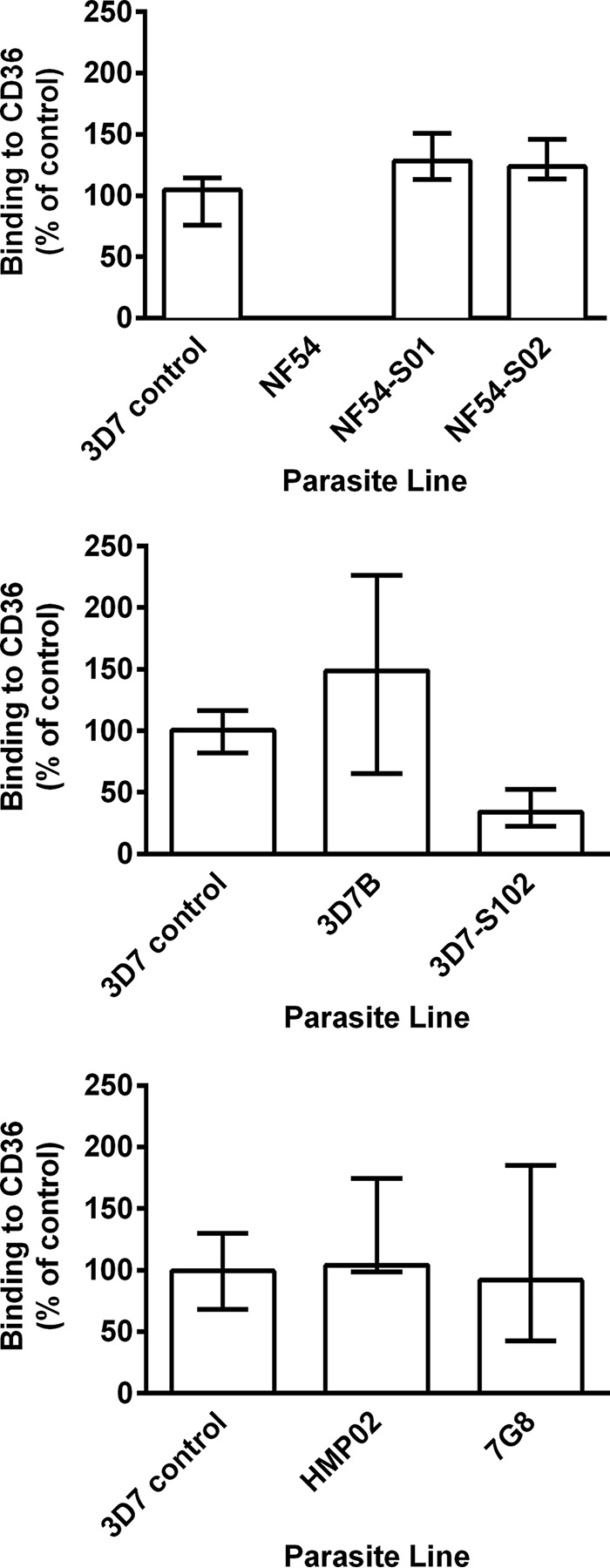
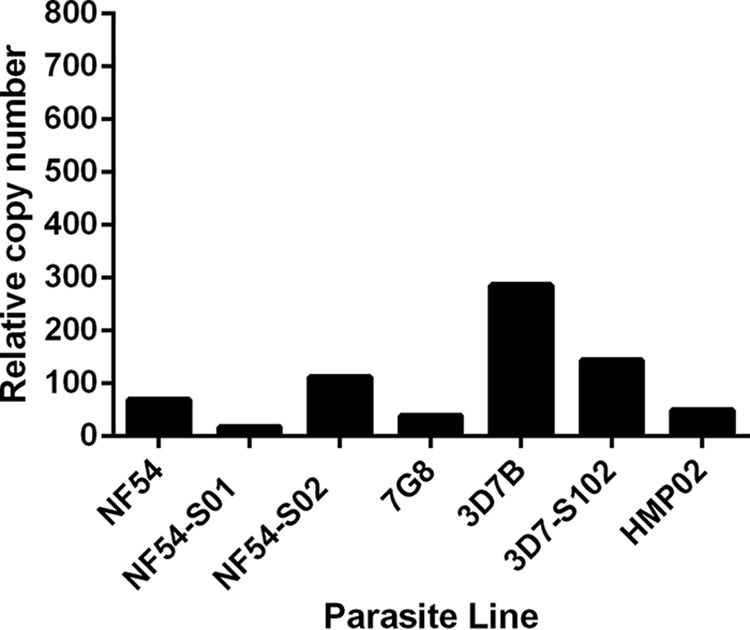
Plasmodium falciparum is the most virulent human malaria parasite because of its ability to cytoadhere in the microvasculature. Nonhuman primate studies demonstrated relationships among knob expression, cytoadherence, and infectivity. This has not been examined in humans. Cultured clinical-grade P. falciparum parasites (NF54, 7G8, and 3D7B) and ex vivo-derived cell banks were characterized. Knob and knob-associated histidine-rich protein expression, CD36 adhesion, and antibody recognition of parasitized erythrocytes (PEs) were evaluated. Parasites from the cell banks were administered to malaria-naive human volunteers to explore infectivity. For the NF54 and 3D7B cell banks, blood was collected from the study participants for in vitro characterization. All parasites were infective in vivo However, infectivity of NF54 was dramatically reduced. In vitro characterization revealed that unlike other cell bank parasites, NF54 PEs lacked knobs and did not cytoadhere. Recognition of NF54 PEs by immune sera was observed, suggesting P. falciparum erythrocyte membrane protein 1 expression. Subsequent recovery of knob expression and CD36-mediated adhesion were observed in PEs derived from participants infected with NF54. Knobless cell bank parasites have a dramatic reduction in infectivity and the ability to adhere to CD36. Subsequent infection of malaria-naive volunteers restored knob expression and CD36-mediated cytoadherence, thereby showing that the human environment can modulate virulence.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
References
-
- Hanson J, Lam SW, Mahanta KC, Pattnaik R, Alam S, Mohanty S, Hasan MU, Hossain A, Charunwatthana P, Chotivanich K, Maude RJ, Kingston H, Day NP, Mishra S, White NJ, Dondorp AM. 2012. Relative contributions of macrovascular and microvascular dysfunction to disease severity in falciparum malaria. J Infect Dis 206:571–579. doi:10.1093/infdis/jis400. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
