

Genetic dissection of Flaviviridae host factors through genome-scale CRISPR screens
- PMID: 27383987
- PMCID: PMC4964798
- DOI: 10.1038/nature18631
Genetic dissection of Flaviviridae host factors through genome-scale CRISPR screens
Abstract
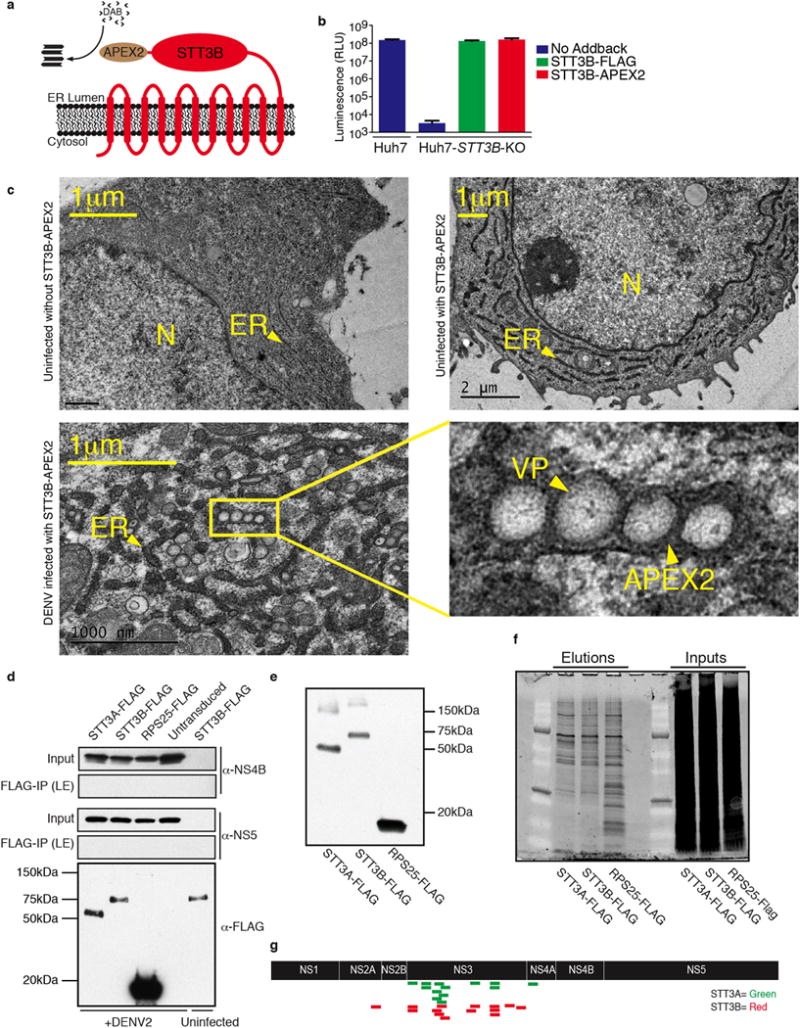
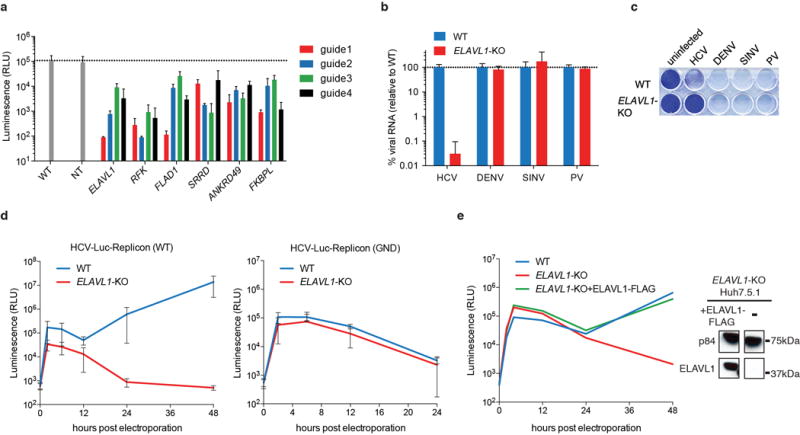
The Flaviviridae are a family of viruses that cause severe human diseases. For example, dengue virus (DENV) is a rapidly emerging pathogen causing an estimated 100 million symptomatic infections annually worldwide. No approved antivirals are available to date and clinical trials with a tetravalent dengue vaccine showed disappointingly low protection rates. Hepatitis C virus (HCV) also remains a major medical problem, with 160 million chronically infected patients worldwide and only expensive treatments available. Despite distinct differences in their pathogenesis and modes of transmission, the two viruses share common replication strategies. A detailed understanding of the host functions that determine viral infection is lacking. Here we use a pooled CRISPR genetic screening strategy to comprehensively dissect host factors required for these two highly important Flaviviridae members. For DENV, we identified endoplasmic-reticulum (ER)-associated multi-protein complexes involved in signal sequence recognition, N-linked glycosylation and ER-associated degradation. DENV replication was nearly completely abrogated in cells deficient in the oligosaccharyltransferase (OST) complex. Mechanistic studies pinpointed viral RNA replication and not entry or translation as the crucial step requiring the OST complex. Moreover, we show that viral non-structural proteins bind to the OST complex. The identified ER-associated protein complexes were also important for infection by other mosquito-borne flaviviruses including Zika virus, an emerging pathogen causing severe birth defects. By contrast, the most significant genes identified in the HCV screen were distinct and included viral receptors, RNA-binding proteins and enzymes involved in metabolism. We found an unexpected link between intracellular flavin adenine dinucleotide (FAD) levels and HCV replication. This study shows notable divergence in host-depenency factors between DENV and HCV, and illuminates new host targets for antiviral therapy.
Conflict of interest statement
The authors declare no competing financial interest.
Figures












References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
