

Chromosome Duplication in Saccharomyces cerevisiae
- PMID: 27384026
- PMCID: PMC4937469
- DOI: 10.1534/genetics.115.186452
Chromosome Duplication in Saccharomyces cerevisiae
Abstract
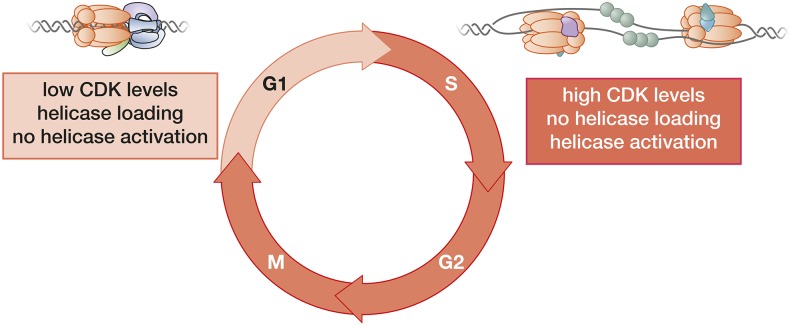
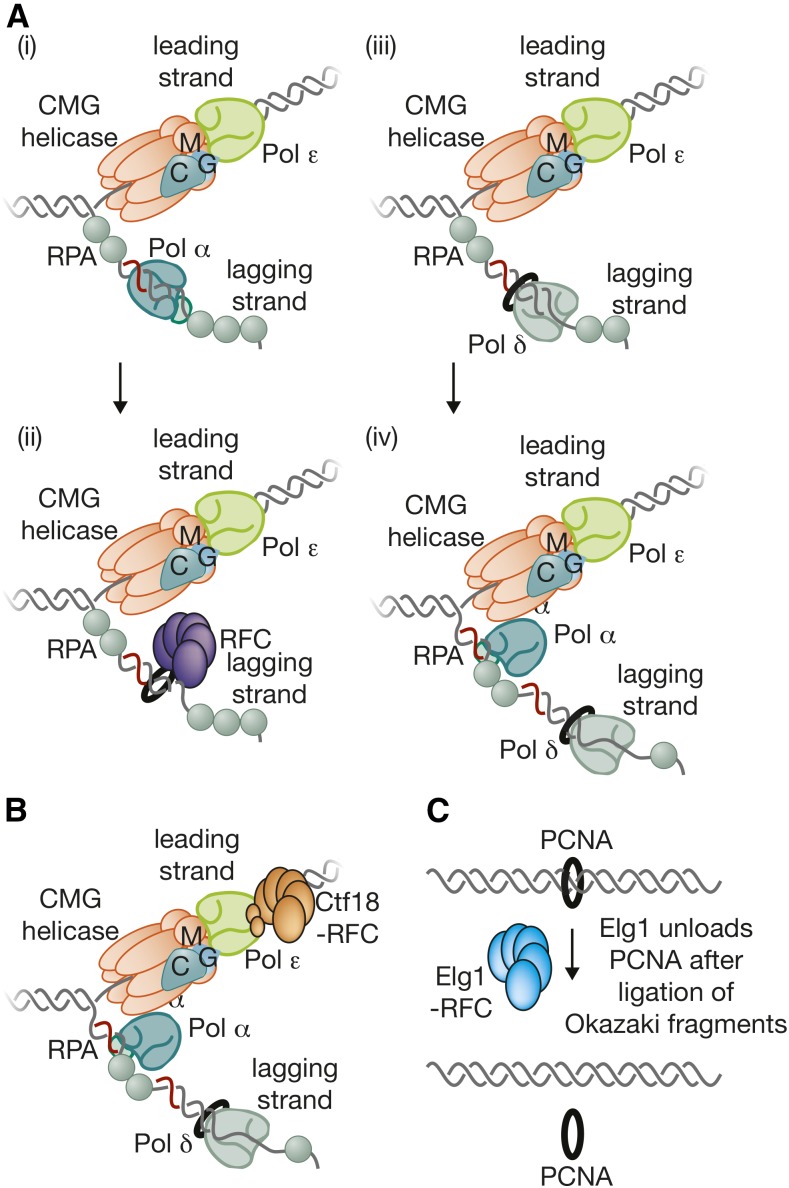
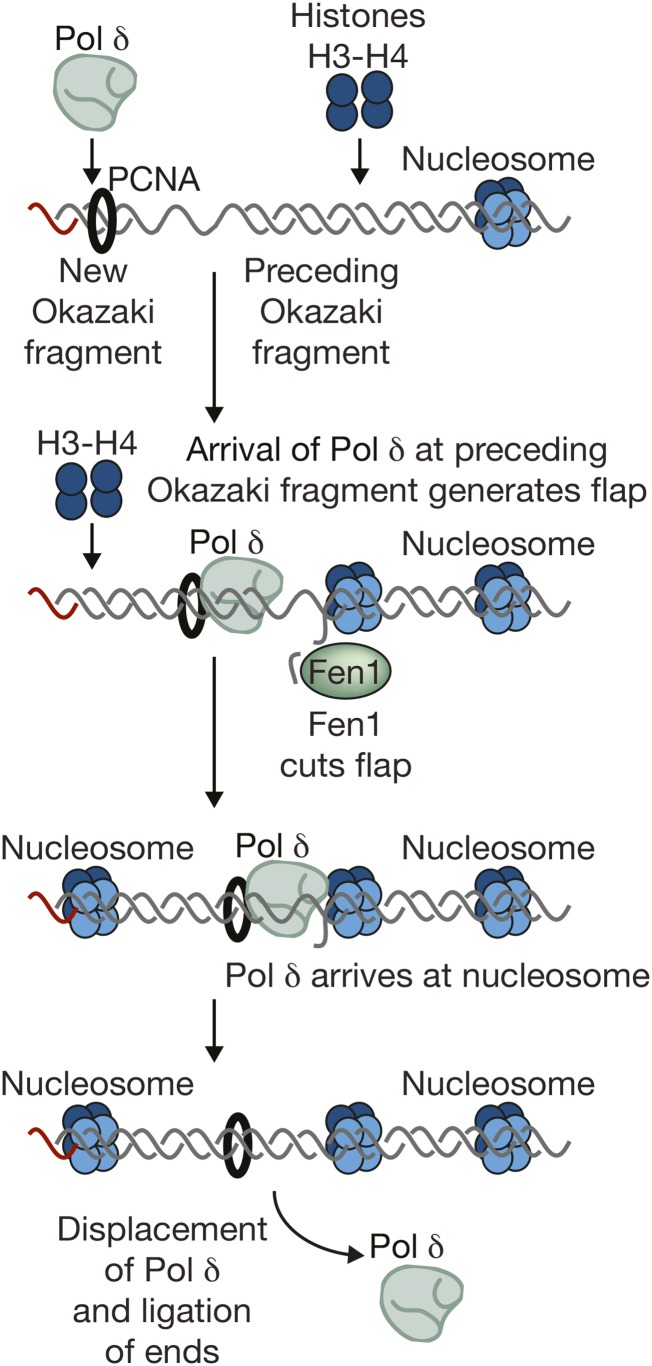
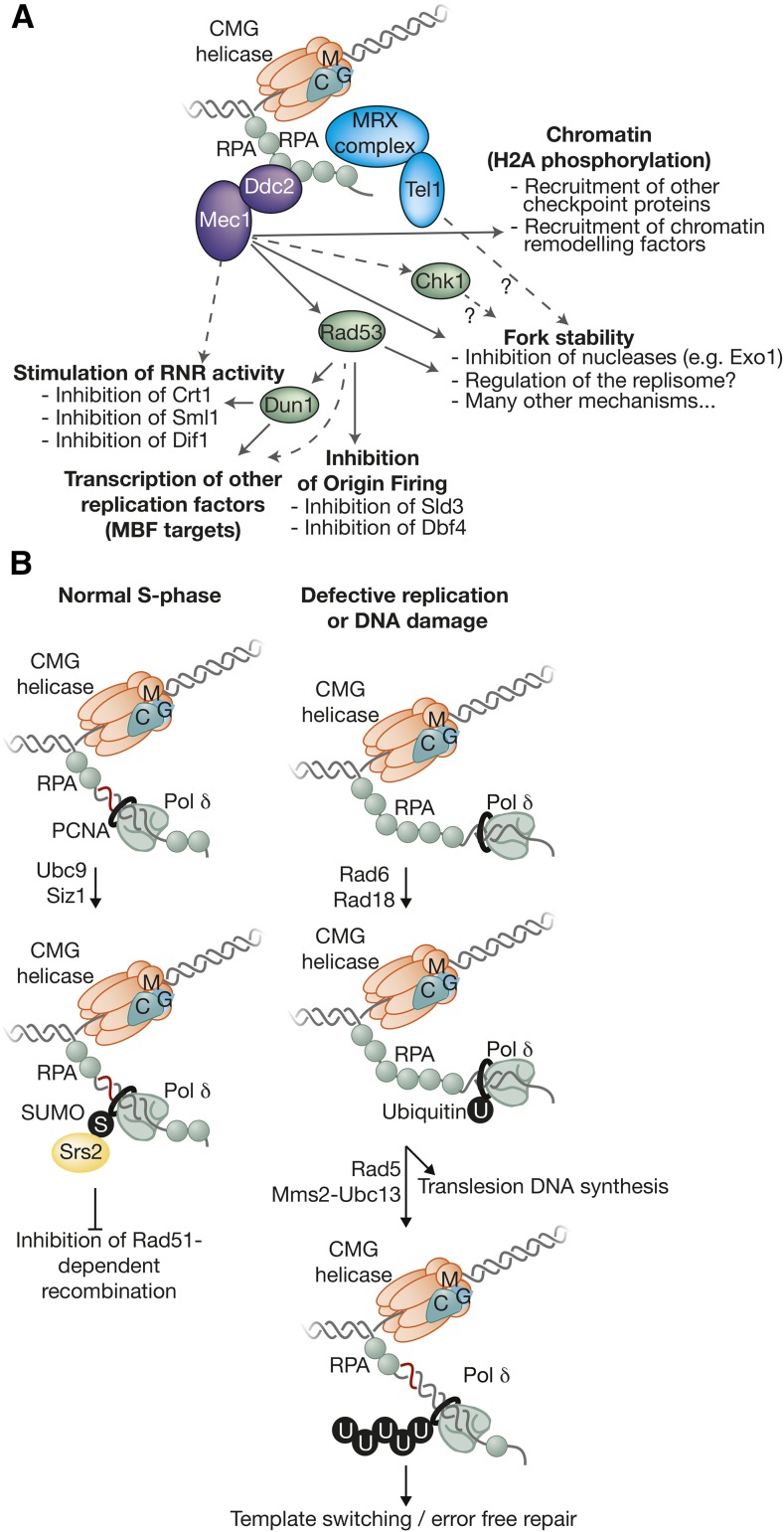
The accurate and complete replication of genomic DNA is essential for all life. In eukaryotic cells, the assembly of the multi-enzyme replisomes that perform replication is divided into stages that occur at distinct phases of the cell cycle. Replicative DNA helicases are loaded around origins of DNA replication exclusively during G1 phase. The loaded helicases are then activated during S phase and associate with the replicative DNA polymerases and other accessory proteins. The function of the resulting replisomes is monitored by checkpoint proteins that protect arrested replisomes and inhibit new initiation when replication is inhibited. The replisome also coordinates nucleosome disassembly, assembly, and the establishment of sister chromatid cohesion. Finally, when two replisomes converge they are disassembled. Studies in Saccharomyces cerevisiae have led the way in our understanding of these processes. Here, we review our increasingly molecular understanding of these events and their regulation.
Keywords: DNA replication; YeastBook; cell cycle; chromatin; chromosome duplication; genome stability.
Copyright © 2016 by the Genetics Society of America.
Figures








References
-
- Alabert C., Groth A., 2012. Chromatin replication and epigenome maintenance. Nat. Rev. Mol. Cell Biol. 13: 153–167. - PubMed
-
- Albert I., Mavrich T. N., Tomsho L. P., Qi J., Zanton S. J., et al. 2007. Translational and rotational settings of H2A.Z nucleosomes across the Saccharomyces cerevisiae genome. Nature 446: 572–576. - PubMed
-
- Alcasabas A. A., Osborn A. J., Bachant J., Hu F., Werler P. J., et al. 2001. Mrc1 transduces signals of DNA replication stress to activate Rad53. Nat. Cell Biol. 3: 958–965. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
