

The "Ram Effect": A "Non-Classical" Mechanism for Inducing LH Surges in Sheep
- PMID: 27384667
- PMCID: PMC4934854
- DOI: 10.1371/journal.pone.0158530
The "Ram Effect": A "Non-Classical" Mechanism for Inducing LH Surges in Sheep
Abstract
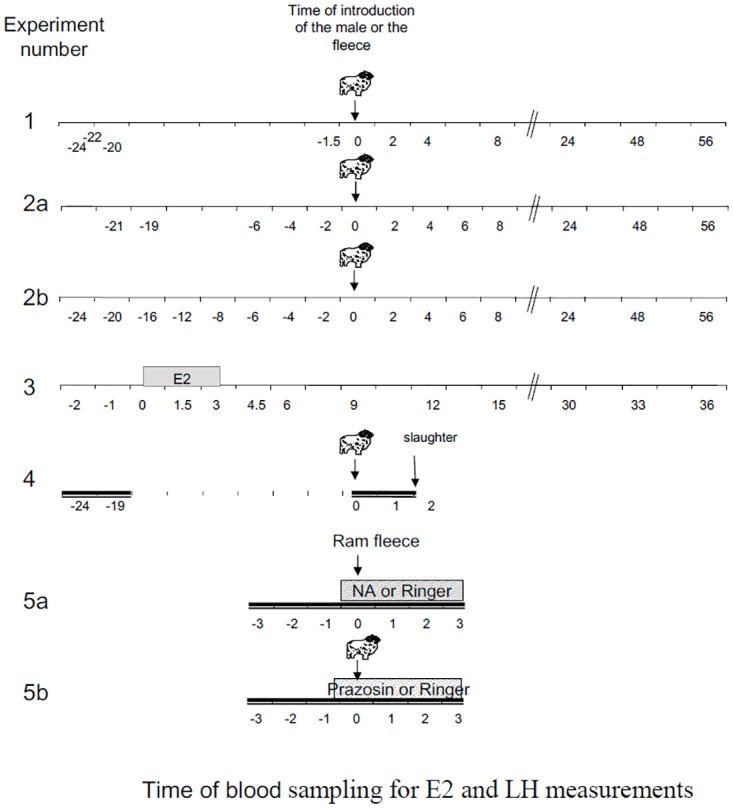
During spring sheep do not normally ovulate but exposure to a ram can induce ovulation. In some ewes an LH surge is induced immediately after exposure to a ram thus raising questions about the control of this precocious LH surge. Our first aim was to determine the plasma concentrations of oestradiol (E2) E2 in anoestrous ewes before and after the "ram effect" in ewes that had a "precocious" LH surge (starting within 6 hours), a "normal" surge (between 6 and 28h) and "late» surge (not detected by 56h). In another experiment we tested if a small increase in circulating E2 could induce an LH surge in anoestrus ewes. The concentration of E2 significantly was not different at the time of ram introduction among ewes with the three types of LH surge. "Precocious" LH surges were not preceded by a large increase in E2 unlike "normal" surges and small elevations of circulating E2 alone were unable to induce LH surges. These results show that the "precocious" LH surge was not the result of E2 positive feedback. Our second aim was to test if noradrenaline (NA) is involved in the LH response to the "ram effect". Using double labelling for Fos and tyrosine hydroxylase (TH) we showed that exposure of anoestrous ewes to a ram induced a higher density of cells positive for both in the A1 nucleus and the Locus Coeruleus complex compared to unstimulated controls. Finally, the administration by retrodialysis into the preoptic area, of NA increased the proportion of ewes with an LH response to ram odor whereas treatment with the α1 antagonist Prazosin decreased the LH pulse frequency and amplitude induced by a sexually active ram. Collectively these results suggest that in anoestrous ewes NA is involved in ram-induced LH secretion as observed in other induced ovulators.
Conflict of interest statement
Figures







References
-
- Corker CS, Naftolin F, Exley D. Interrelationship between plasma luteinizing hormone and oestradiol in the human menstrual cycle. Nature 1969; 222:1063 - PubMed
-
- Smith MS, Freeman ME, Neill JD. The control of progesterone secretion during the estrous cycle and early pseudopregnancy in the rat: prolactin, gonadotropin and steroid levels associated with rescue of the corpus luteum of pseudopregnancy. Endocrinology 1975; 96: 219–226. - PubMed
-
- Hauger RL, Karsch FJ, Foster DL. A new concept for control of the estrous cycle of the ewe based on the temporal relationships between luteinizing hormone, estradiol and progesterone in peripheral serum and evidence that progesterone inhibits tonic LH secretion. Endocrinology. 1977; 101:807–817. - PubMed
-
- Karsch FJ, Weick RF, Butler WR, Dierschke DJ, Krey LC, Weiss G et al. Induced LH surges in the rhesus monkey: strength-duration characteristics of the estrogen stimulus. Endocrinology. 1973; 92: 1740–1747. - PubMed
-
- Karsch FJ, Foster DL, Legan SJ, Ryan KD, Peter GK. Control of the preovulatory endocrine events in the ewe: interrelationship of estradiol, progesterone, and luteinizing hormone. Endocrinology. 1979; 105: 421–426. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
