

IMP2 axonal localization, RNA interactome, and function in the development of axon trajectories
- PMID: 27385015
- PMCID: PMC5004904
- DOI: 10.1242/dev.128348
IMP2 axonal localization, RNA interactome, and function in the development of axon trajectories
Abstract
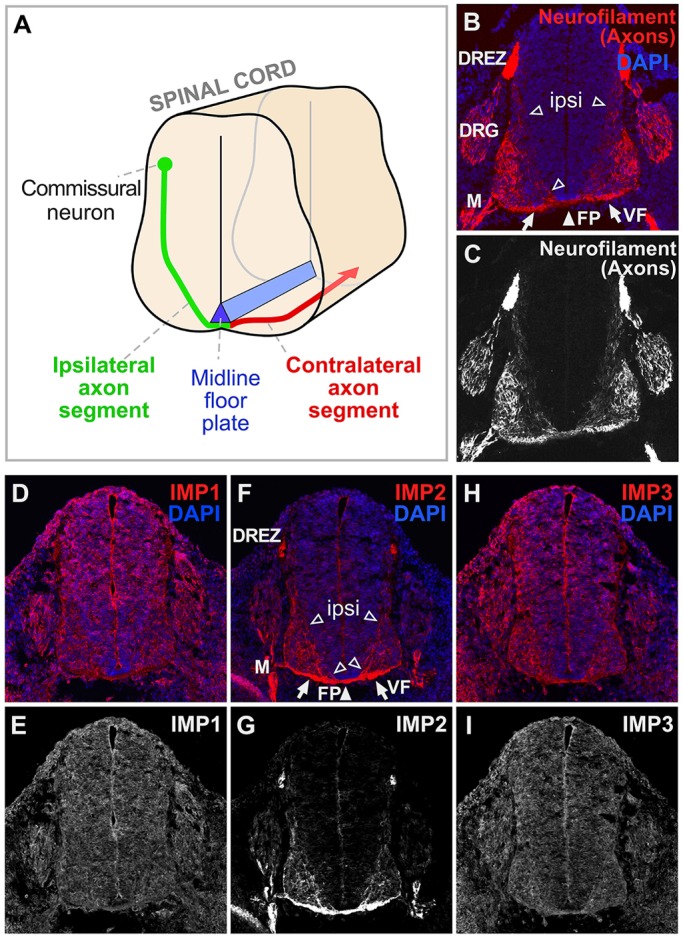
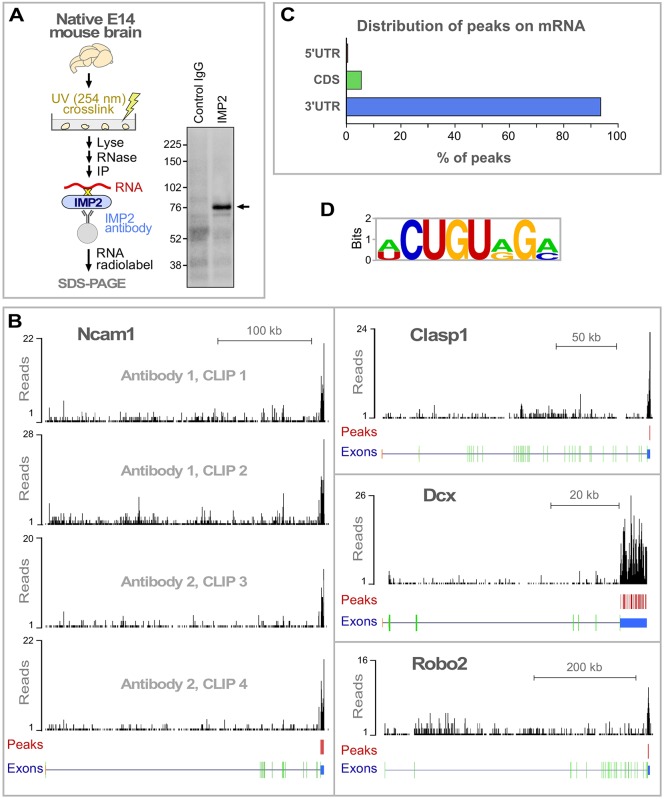
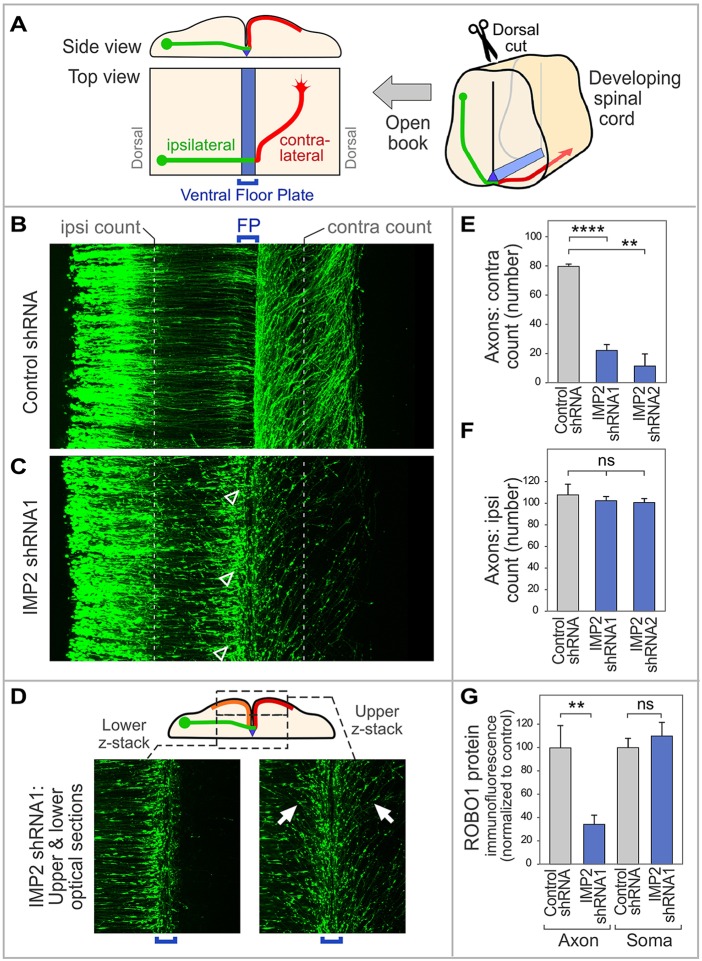
RNA-based regulatory mechanisms play important roles in the development and plasticity of neural circuits and neurological disease. Developing axons provide a model well suited to the study of RNA-based regulation, and contain specific subsets of mRNAs that are locally translated and have roles in axon pathfinding. However, the RNA-binding proteins involved in axon pathfinding, and their corresponding mRNA targets, are still largely unknown. Here we find that the RNA-binding protein IMP2 (Igf2bp2) is strikingly enriched in developing axon tracts, including in spinal commissural axons. We used the HITS-CLIP approach to perform a genome-wide identification of RNAs that interact directly with IMP2 in the native context of developing mouse brain. This IMP2 interactome was highly enriched for mRNA targets related to axon guidance. Accordingly, IMP2 knockdown in the developing spinal cord led to strong defects in commissural axon trajectories at the midline intermediate target. These results reveal a highly distinctive axonal enrichment of IMP2, show that it interacts with a network of axon guidance-related mRNAs, and reveal that it is required for normal axon pathfinding during vertebrate development.
Keywords: Axon guidance; IMP2; Igf2bp2; RNA interactome; RNA-binding protein.
© 2016. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
