

A Critical Role for CD200R Signaling in Limiting the Growth and Metastasis of CD200+ Melanoma
- PMID: 27385779
- PMCID: PMC4975953
- DOI: 10.4049/jimmunol.1600052
A Critical Role for CD200R Signaling in Limiting the Growth and Metastasis of CD200+ Melanoma
Abstract
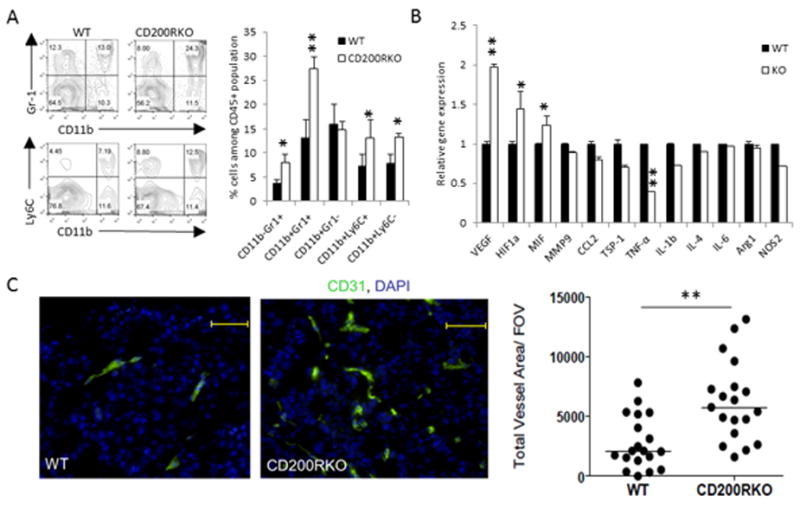
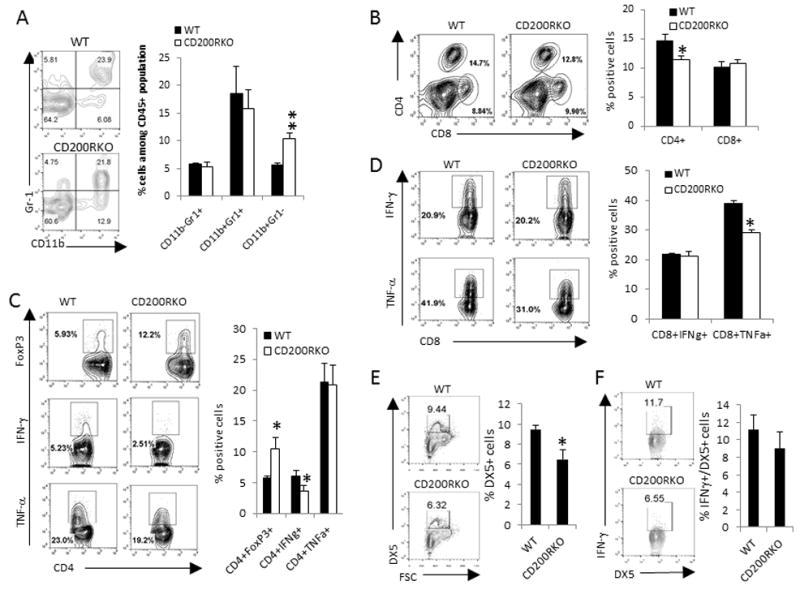
CD200 is a cell surface glycoprotein that functions through engaging CD200R on cells of the myeloid lineage and inhibits their functions. Expression of CD200 was implicated in a variety of human cancer cells, including melanoma cells; however, its roles in tumor growth and immunity are not clearly understood. In this study, we used CD200R-deficient mice and the B16 tumor model to evaluate this issue. We found that CD200R-deficient mice exhibited accelerated growth of CD200(+), but not CD200(-), B16 tumors. Strikingly, CD200R-deficient mice receiving CD200(+) B16 cells i.v. exhibited massive tumor growth in multiple organs, including liver, lung, kidney, and peritoneal cavity, whereas the growth of the same tumors in wild-type mice was limited. CD200(+) tumors grown in CD200R-deficient mice contained higher numbers of CD11b(+)Ly6C(+) myeloid cells, exhibited increased expression of VEGF and HIF1α genes with increased angiogenesis, and showed significantly reduced infiltration of CD4(+) and CD8(+) T cells, presumably as the result of reduced expression of T cell chemokines, such as CXCL9 and CXCL16. The liver from CD200R-deficient mice, under metastatic growth of CD200(+) tumors, contained significantly increased numbers of CD11b(+)Gr1(-) myeloid cells and Foxp3(+) regulatory T cells and reduced numbers of NK cells. Liver T cells also had a reduced capacity to produce IFN-γ or TNF-α. Taken together, we revealed a critical role for CD200R signaling in limiting the growth and metastasis of CD200(+) tumors. Thus, targeting CD200R signaling may potentially interfere with the metastatic growth of CD200(+) tumors, like melanoma.
Copyright © 2016 by The American Association of Immunologists, Inc.
Figures





References
-
- Mantovani A, Allavena P, Sica A, Balkwill F. Cancer-related inflammation. Nature. 2008;454:436–444. - PubMed
-
- Yang L, DeBusk LM, Fukuda K, Fingleton B, Green-Jarvis B, Shyr Y, Matrisian LM, Carbone DP, Lin PC. Expansion of myeloid immune suppressor Gr+CD11b+ cells in tumor-bearing host directly promotes tumor angiogenesis. Cancer Cell. 2004;6:409–421. - PubMed
-
- Murdoch C, Muthana M, Coffelt SB, Lewis CE. The role of myeloid cells in the promotion of tumour angiogenesis. Nat Rev Cancer. 2008;8:618–631. - PubMed
-
- Mantovani A, Schioppa T, Porta C, Allavena P, Sica A. Role of tumor-associated macrophages in tumor progression and invasion. Cancer Metastasis Rev. 2006;25:315–322. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
