

Novel Monoclonal Antibodies for Studies of Human and Rhesus Macaque Secretory Component and Human J-Chain
- PMID: 27386924
- PMCID: PMC5003010
- DOI: 10.1089/mab.2016.0014
Novel Monoclonal Antibodies for Studies of Human and Rhesus Macaque Secretory Component and Human J-Chain
Abstract
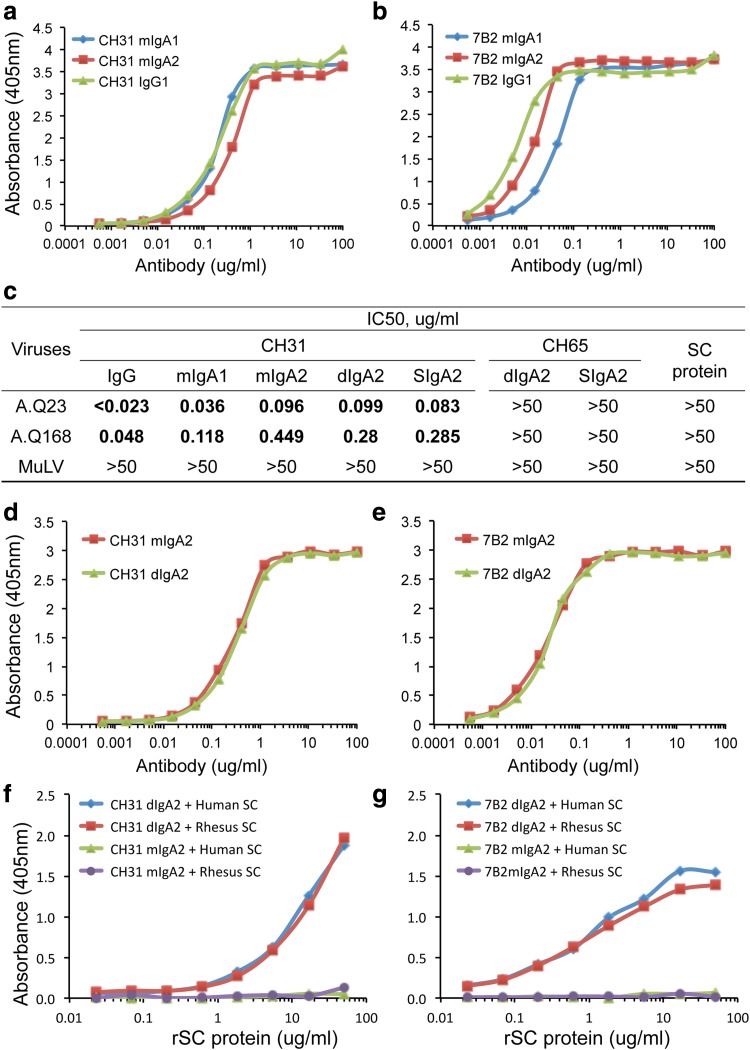
Immunoglobulin A (IgA) antibodies exist in monomeric, dimeric, and secretory forms. Dimerization of IgA depends on a 15-kD polypeptide termed "joining (J) chain," which is also part of the binding site for an epithelial glycoprotein called "secretory component (SC)," whether this after apical cleavage on secretory epithelia is ligand bound in secretory IgA (SIgA) or in a free form. Uncleaved membrane SC, also called the "polymeric Ig receptor," is thus crucial for transcytotic export of SIgA to mucosal surfaces, where it interacts with and modulates commensal bacteria and mediates protective immune responses against exogenous pathogens. To evaluate different forms of IgA, we have produced mouse monoclonal antibodies (MAbs) against human J-chain and free SC. We found that J-chain MAb 9A8 and SC MAb 9H7 identified human dimeric IgA and SIgA in enzyme-linked immunoassay and western blot analysis, as well as functioning in immunohistochemistry to identify cytoplasmic IgA of intestinal lamina propria plasmablasts/plasma cells and crypt epithelium of distal human intestine. Finally, we demonstrated that SC MAb 9H7 cross-reacted with rhesus macaque SIgA. These novel reagents should be of use in the study of the biology of various forms of IgA in humans and SIgA in macaques, as well as in monitoring the production and/or isolation of these forms of IgA.
Figures




References
-
- Brandtzaeg P, Kiyono H, Pabst R, and Russell MW: Terminology: Nomenclature of mucosa-associated lymphoid tissue. Mucosal Immunol 2008;1:31–37 - PubMed
-
- Fagarasan S, and Honjo T: Intestinal IgA synthesis: Regulation of front-line body defences. Nat Rev Immunol 2003;3:63–72 - PubMed
-
- Fernandez MI, Pedron T, Tournebize R, Olivo-Marin JC, Sansonetti PJ, and Phalipon A: Anti-inflammatory role for intracellular dimeric immunoglobulin A by neutralization of lipopolysaccharide in epithelial cells. Immunity 2003;18:739–749 - PubMed
-
- Macpherson AJ, and Uhr T: Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science 2004;303:1662–1665 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
