

Widespread parainflammation in human cancer
- PMID: 27386949
- PMCID: PMC4937599
- DOI: 10.1186/s13059-016-0995-z
Widespread parainflammation in human cancer
Abstract
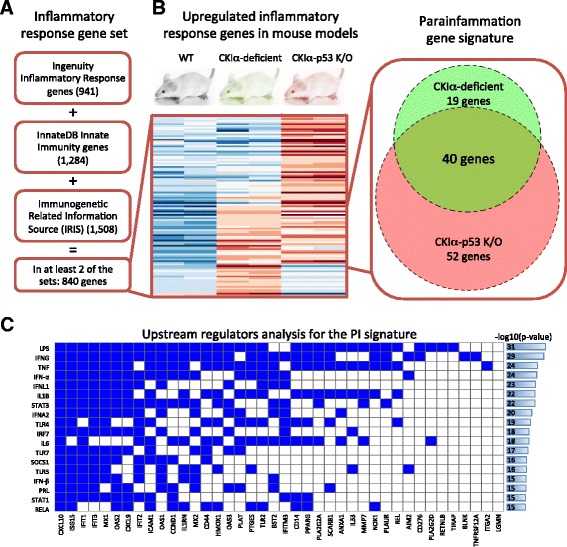
Background: Chronic inflammation has been recognized as one of the hallmarks of cancer. We recently showed that parainflammation, a unique variant of inflammation between homeostasis and chronic inflammation, strongly promotes mouse gut tumorigenesis upon p53 loss. Here we explore the prevalence of parainflammation in human cancer and determine its relationship to certain molecular and clinical parameters affecting treatment and prognosis.
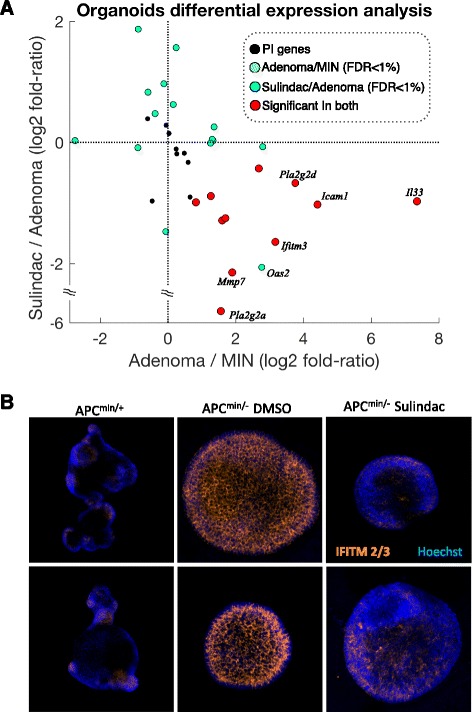
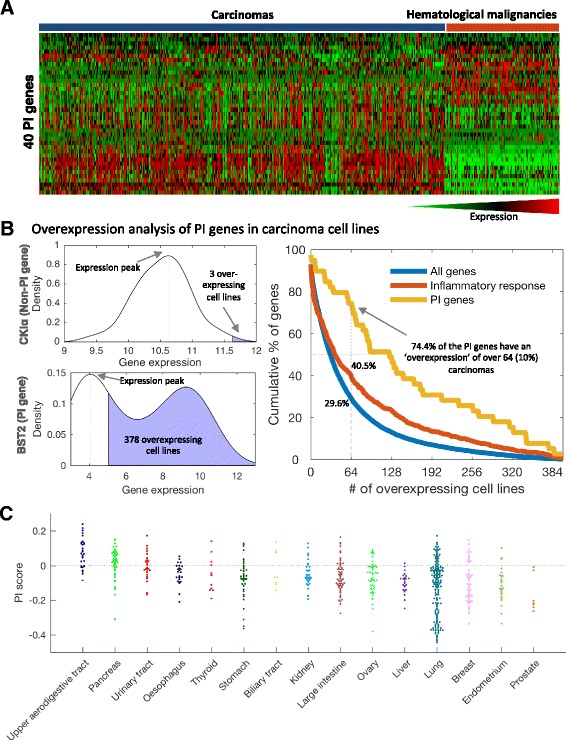
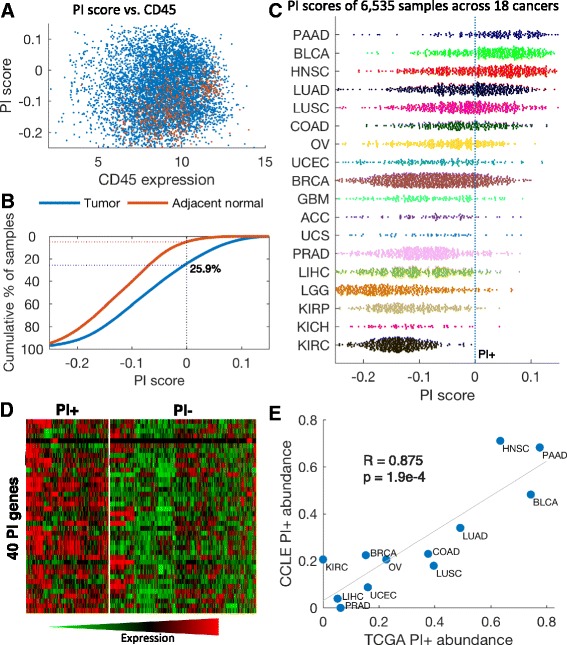
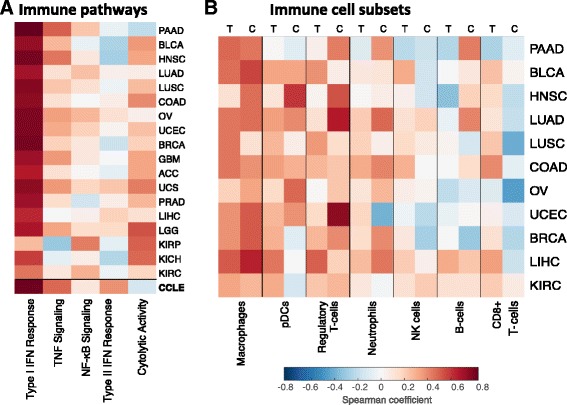
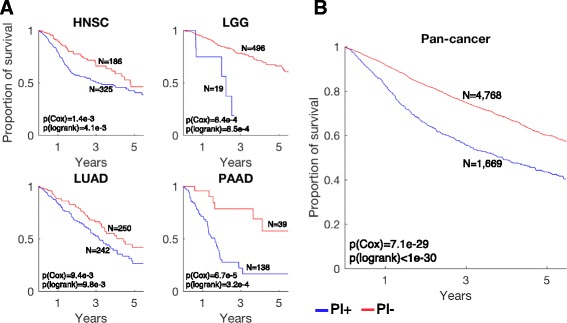
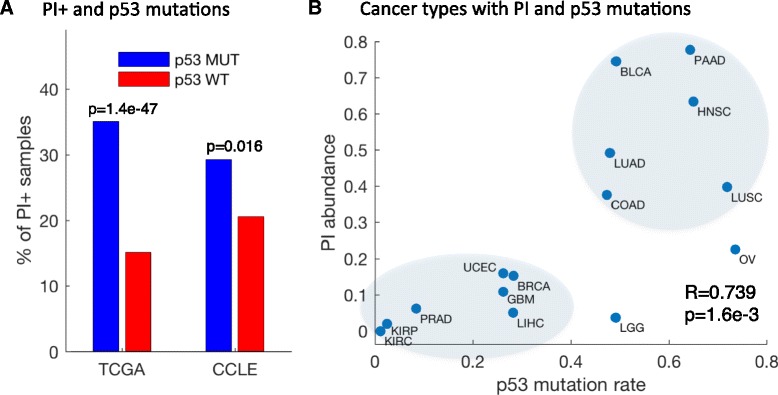
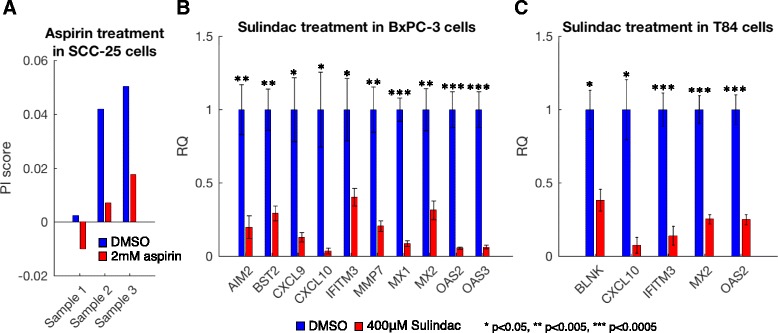
Results: We generated a transcriptome signature to identify parainflammation in many primary human tumors and carcinoma cell lines as distinct from their normal tissue counterparts and the tumor microenvironment and show that parainflammation-positive tumors are enriched for p53 mutations and associated with poor prognosis. Non-steroidal anti-inflammatory drug (NSAID) treatment suppresses parainflammation in both murine and human cancers, possibly explaining a protective effect of NSAIDs against cancer.
Conclusions: We conclude that parainflammation, a low-grade form of inflammation, is widely prevalent in human cancer, particularly in cancer types commonly harboring p53 mutations. Our data suggest that parainflammation may be a driver for p53 mutagenesis and a guide for cancer prevention by NSAID treatment.
Keywords: Cancer prevention; Genomics; Inflammation; NSAID treatment; Parainflammation; p53 mutations.
Figures
References
-
- Elyada E, Pribluda A, Goldstein RE, Morgenstern Y, Brachya G, Cojocaru G, Snir-Alkalay I, Burstain I, Haffner-Krausz R, Jung S, et al. CKIalpha ablation highlights a critical role for p53 in invasiveness control. Nature. 2011;470:409–13. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
