

AP1S3 Mutations Cause Skin Autoinflammation by Disrupting Keratinocyte Autophagy and Up-Regulating IL-36 Production
- PMID: 27388993
- PMCID: PMC5070969
- DOI: 10.1016/j.jid.2016.06.618
AP1S3 Mutations Cause Skin Autoinflammation by Disrupting Keratinocyte Autophagy and Up-Regulating IL-36 Production
Abstract
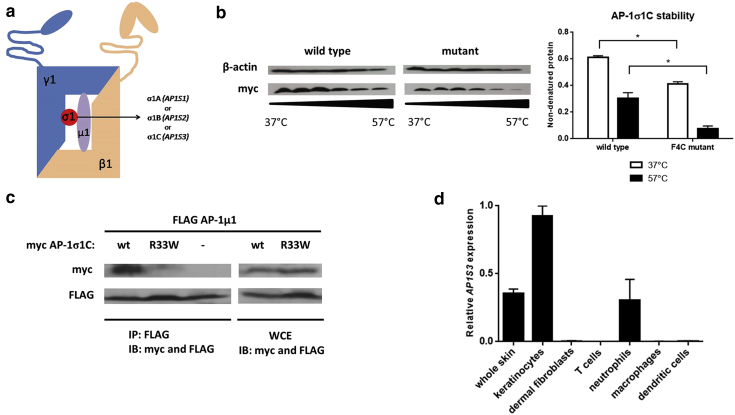
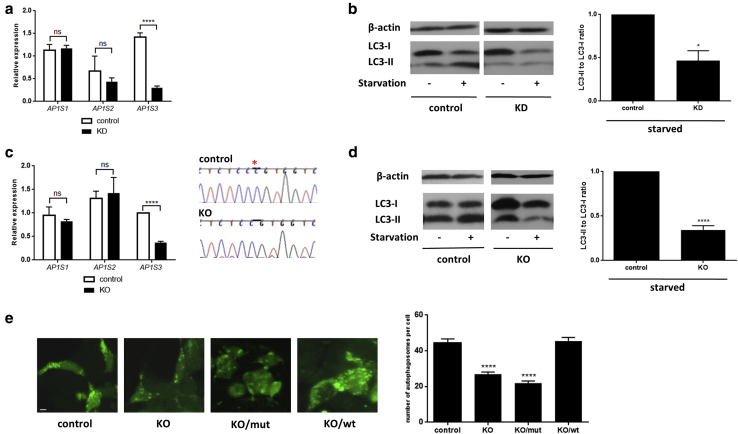
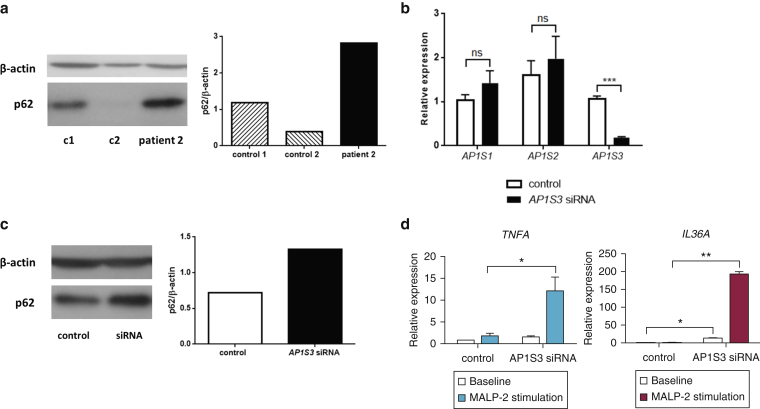
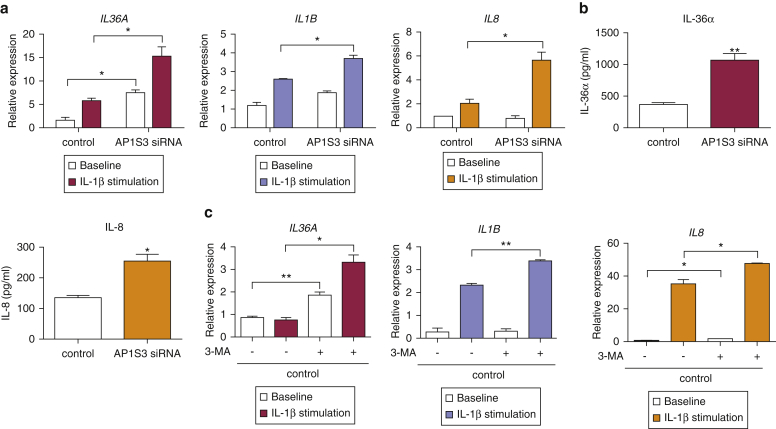
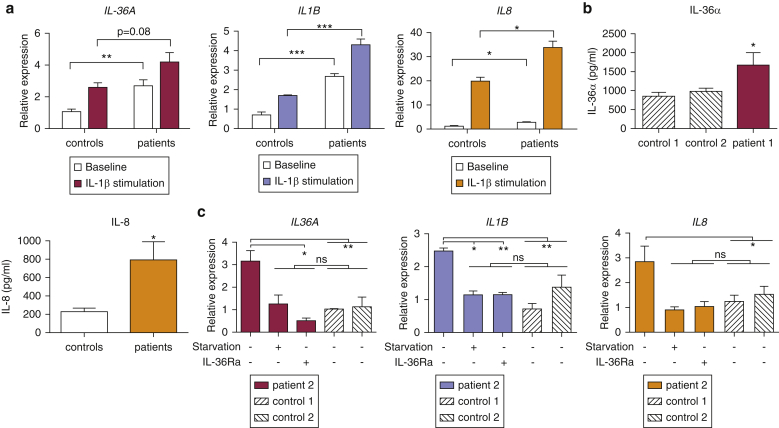
Prominent skin involvement is a defining characteristic of autoinflammatory disorders caused by abnormal IL-1 signaling. However, the pathways and cell types that drive cutaneous autoinflammatory features remain poorly understood. We sought to address this issue by investigating the pathogenesis of pustular psoriasis, a model of autoinflammatory disorders with predominant cutaneous manifestations. We specifically characterized the impact of mutations affecting AP1S3, a disease gene previously identified by our group and validated here in a newly ascertained patient resource. We first showed that AP1S3 expression is distinctively elevated in keratinocytes. Because AP1S3 encodes a protein implicated in autophagosome formation, we next investigated the effects of gene silencing on this pathway. We found that AP1S3 knockout disrupts keratinocyte autophagy, causing abnormal accumulation of p62, an adaptor protein mediating NF-κB activation. We showed that as a consequence, AP1S3-deficient cells up-regulate IL-1 signaling and overexpress IL-36α, a cytokine that is emerging as an important mediator of skin inflammation. These abnormal immune profiles were recapitulated by pharmacological inhibition of autophagy and verified in patient keratinocytes, where they were reversed by IL-36 blockade. These findings show that keratinocytes play a key role in skin autoinflammation and identify autophagy modulation of IL-36 signaling as a therapeutic target.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Aasen T., Izpisua Belmonte J.C. Isolation and cultivation of human keratinocytes from skin or plucked hair for the generation of induced pluripotent stem cells. Nat Protoc. 2010;5:371–382. - PubMed
-
- Aksentijevich I., Kastner D.L. Genetics of monogenic autoinflammatory diseases: past successes, future challenges. Nat Rev Rheumatol. 2011;7:469–478. - PubMed
-
- Beer H.D., Contassot E., French L.E. The inflammasomes in autoinflammatory diseases with skin involvement. J Invest Dermatol. 2014;134:1805–1810. - PubMed
-
- Berki D.M., Liu L., Choon S.E., David Burden A., Griffiths C.E., Navarini A.A. Activating CARD14 mutations are associated with generalized pustular psoriasis but rarely account for familial recurrence in psoriasis vulgaris. J Invest Dermatol. 2015;135:2964–2970. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
