

Characterization of two PEBP genes, SrFT and SrMFT, in thermogenic skunk cabbage (Symplocarpus renifolius)
- PMID: 27389636
- PMCID: PMC4937424
- DOI: 10.1038/srep29440
Characterization of two PEBP genes, SrFT and SrMFT, in thermogenic skunk cabbage (Symplocarpus renifolius)
Abstract
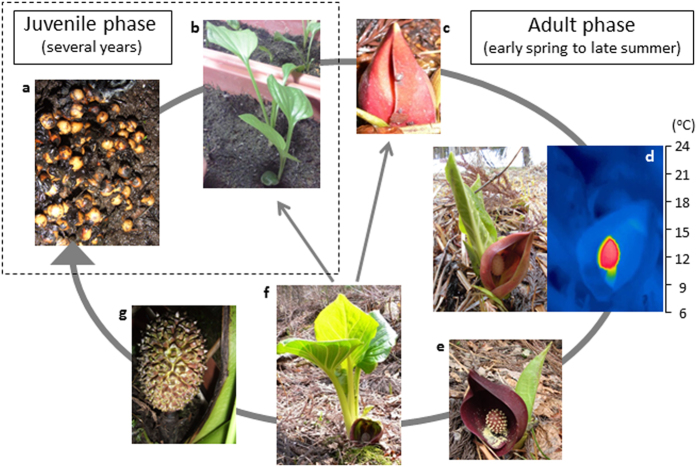
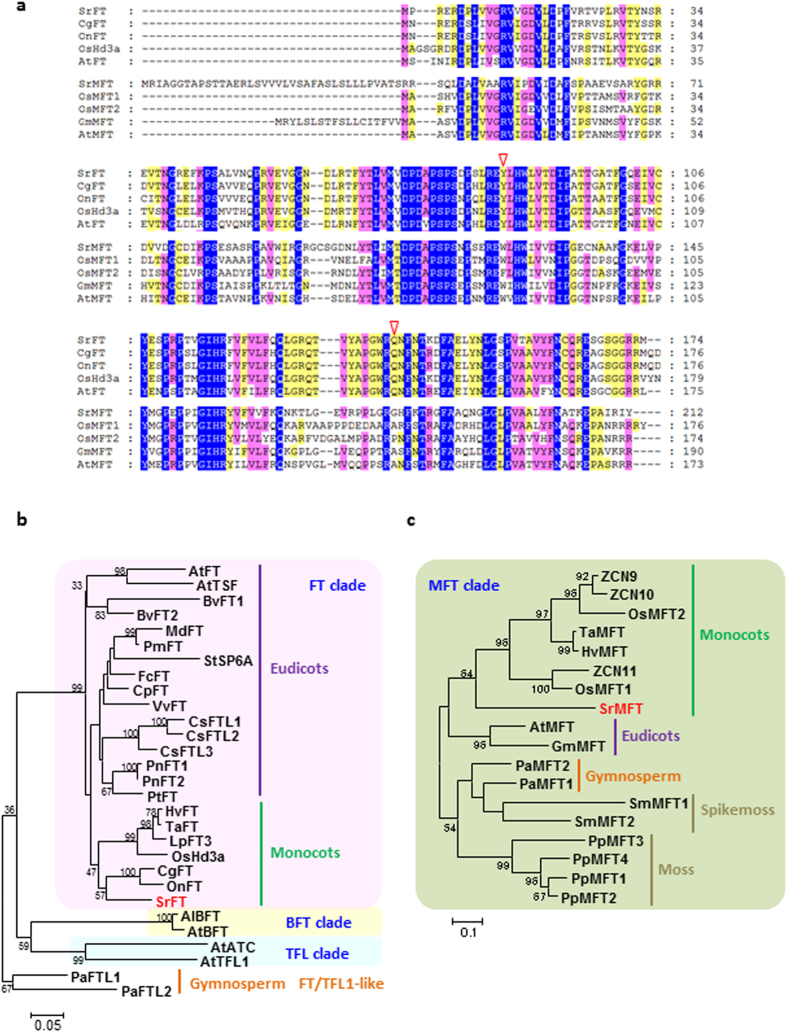
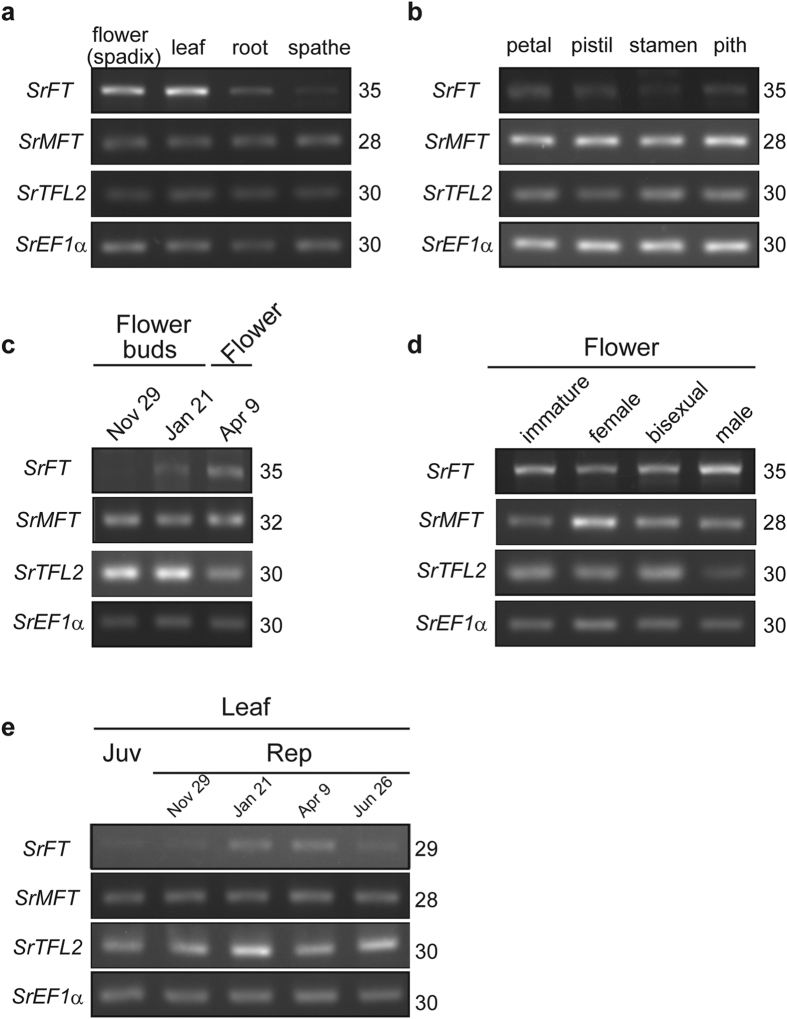
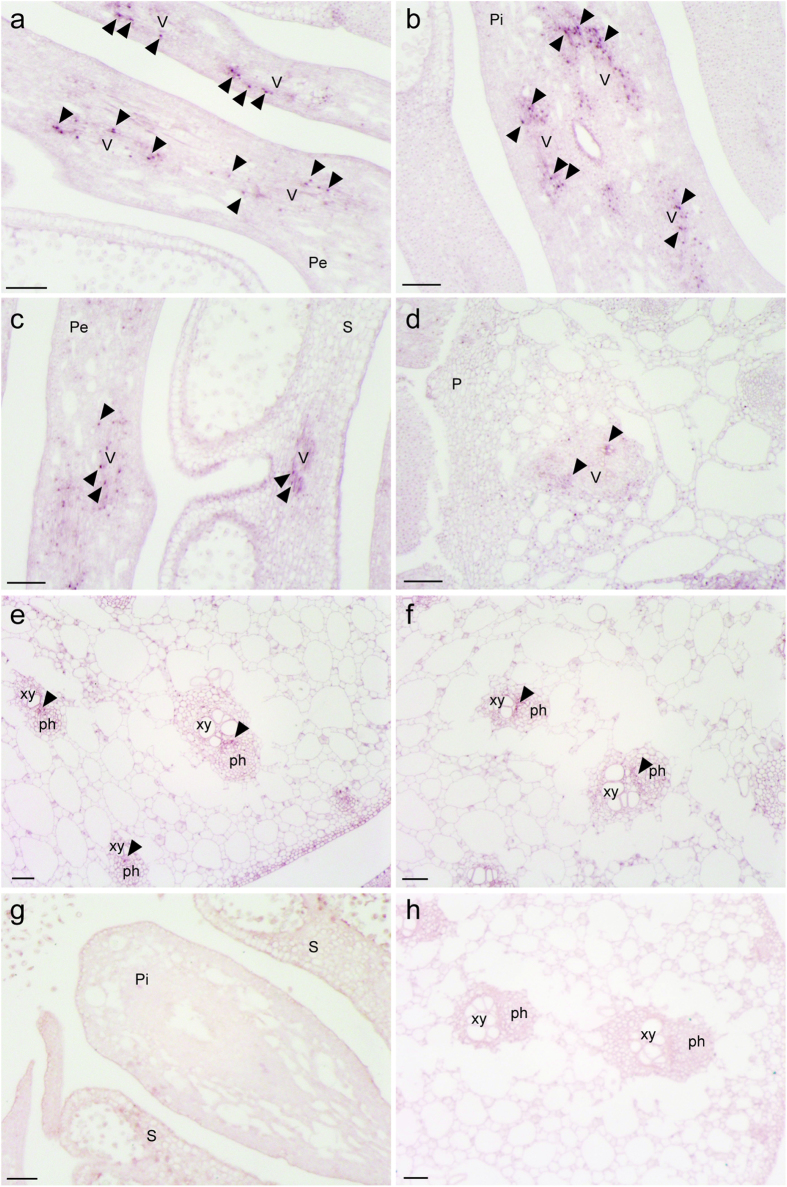
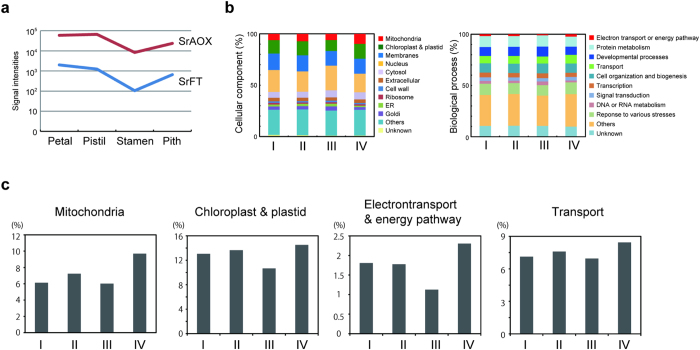
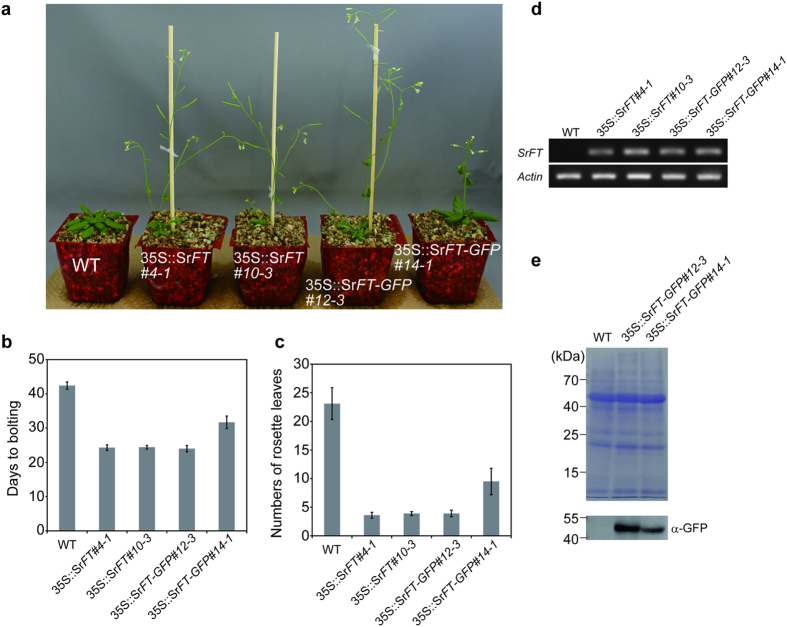
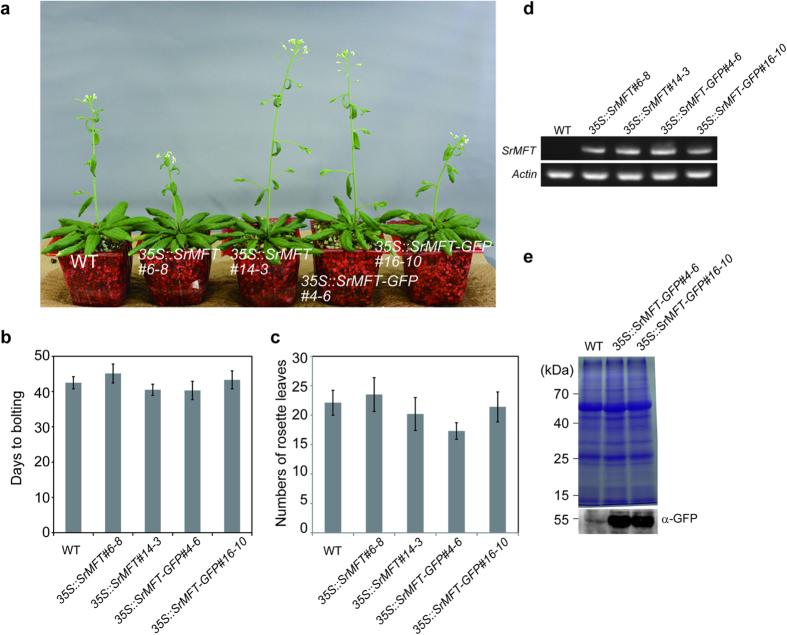
Floral thermogenesis has been found in dozens of primitive seed plants and the reproductive organs in these plants produce heat during anthesis. Thus, characterization of the molecular mechanisms underlying flowering is required to fully understand the role of thermogenesis, but this aspect of thermogenic plant development is largely unknown. In this study, extensive database searches and cloning experiments suggest that thermogenic skunk cabbage (Symplocarpus renifolius), which is a member of the family Araceae, possesses two genes encoding phosphatidyl ethanolamine-binding proteins (PEBP), FLOWERING LOCUS T (SrFT) and MOTHER OF FT AND TFL1 (SrMFT). Functional analyses of SrFT and SrMFT in Arabidopsis indicate that SrFT promotes flowering, whereas SrMFT does not. In S. renifolius, the stage- and tissue-specific expression of SrFT was more evident than that of SrMFT. SrFT was highly expressed in flowers and leaves and was mainly localized in fibrovascular tissues. In addition, microarray analysis revealed that, within floral tissues, SrFT was co-regulated with the genes associated with cellular respiration and mitochondrial function, including ALTERNATIVE OXIDASE gene proposed to play a major role in floral thermogenesis. Taken together, these data suggest that, among the PEBP genes, SrFT plays a role in flowering and floral development in the thermogenic skunk cabbage.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Thioredoxin o-mediated reduction of mitochondrial alternative oxidase in the thermogenic skunk cabbage Symplocarpus renifolius.J Biochem. 2019 Jan 1;165(1):57-65. doi: 10.1093/jb/mvy082. J Biochem. 2019. PMID: 30289493 Free PMC article.
-
Proteins from the FLOWERING LOCUS T-like subclade of the PEBP family act antagonistically to regulate floral initiation in tobacco.Plant J. 2012 Dec;72(6):908-21. doi: 10.1111/j.1365-313X.2012.05125.x. Epub 2012 Oct 15. Plant J. 2012. PMID: 22889438
-
Developmental changes and organelle biogenesis in the reproductive organs of thermogenic skunk cabbage (Symplocarpus renifolius).J Exp Bot. 2009;60(13):3909-22. doi: 10.1093/jxb/erp226. Epub 2009 Jul 29. J Exp Bot. 2009. PMID: 19640927 Free PMC article.
-
Flowering time genes branching out.J Exp Bot. 2024 Jul 23;75(14):4195-4209. doi: 10.1093/jxb/erae112. J Exp Bot. 2024. PMID: 38470076 Free PMC article. Review.
-
PHOSPHATIDYLETHANOLAMINE-BINDING PROTEINS: the conductors of dual reproduction in plants with vegetative storage organs.J Exp Bot. 2021 Apr 2;72(8):2845-2856. doi: 10.1093/jxb/erab064. J Exp Bot. 2021. PMID: 33606013 Review.
Cited by
-
Large-scale analyses of angiosperm Flowering Locus T genes reveal duplication and functional divergence in monocots.Front Plant Sci. 2023 Jan 4;13:1039500. doi: 10.3389/fpls.2022.1039500. eCollection 2022. Front Plant Sci. 2023. PMID: 36684773 Free PMC article.
-
Ubiquitin-Proteasome Dependent Regulation of the GOLDEN2-LIKE 1 Transcription Factor in Response to Plastid Signals.Plant Physiol. 2017 Jan;173(1):524-535. doi: 10.1104/pp.16.01546. Epub 2016 Nov 7. Plant Physiol. 2017. PMID: 27821720 Free PMC article.
-
Establishing an efficient protoplast transient expression system for investigation of floral thermogenesis in aroids.Plant Cell Rep. 2022 Jan;41(1):263-275. doi: 10.1007/s00299-021-02806-1. Epub 2021 Oct 26. Plant Cell Rep. 2022. PMID: 34704119
-
Identification of cotton MOTHER OF FT AND TFL1 homologs, GhMFT1 and GhMFT2, involved in seed germination.PLoS One. 2019 Apr 19;14(4):e0215771. doi: 10.1371/journal.pone.0215771. eCollection 2019. PLoS One. 2019. PMID: 31002698 Free PMC article.
-
Seed-Specific Gene MOTHER of FT and TFL1(MFT) Involved in Embryogenesis, Hormones and Stress Responses in Dimocarpus longan Lour.Int J Mol Sci. 2018 Aug 14;19(8):2403. doi: 10.3390/ijms19082403. Int J Mol Sci. 2018. PMID: 30110985 Free PMC article.
References
-
- Yuan L.-C. et al.. Pollination of Kadsura longipedunculata (Schisandraceae), a monoecious basal angiosperm, by female, pollen-eating Megommata sp. (Cecidomyiidae: Diptera) in China. Biol J Linn Soc 93, 523–536, doi: 10.1111/j.1095-8312.2007.00897.x (2008). - DOI
-
- Ervik F. & Knudsen J. T. Water lilies and scarabs: faithful partners for 100 million years? Biol J Linn Soc 80, 539–543, doi: 10.1046/j.1095-8312.2003.00258.x (2003). - DOI
-
- Seymour R. S., Silberbauer-Gottsberger I. & Gottsberger G. Respiration and temperature patterns in thermogenic flowers of Magnolia ovata under natural conditions in Brazil. Funct Plant Biol 37, 870–878, 10.1071/FP10039 (2010). - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
