

Properties of Neurons in External Globus Pallidus Can Support Optimal Action Selection
- PMID: 27389780
- PMCID: PMC4936724
- DOI: 10.1371/journal.pcbi.1005004
Properties of Neurons in External Globus Pallidus Can Support Optimal Action Selection
Abstract
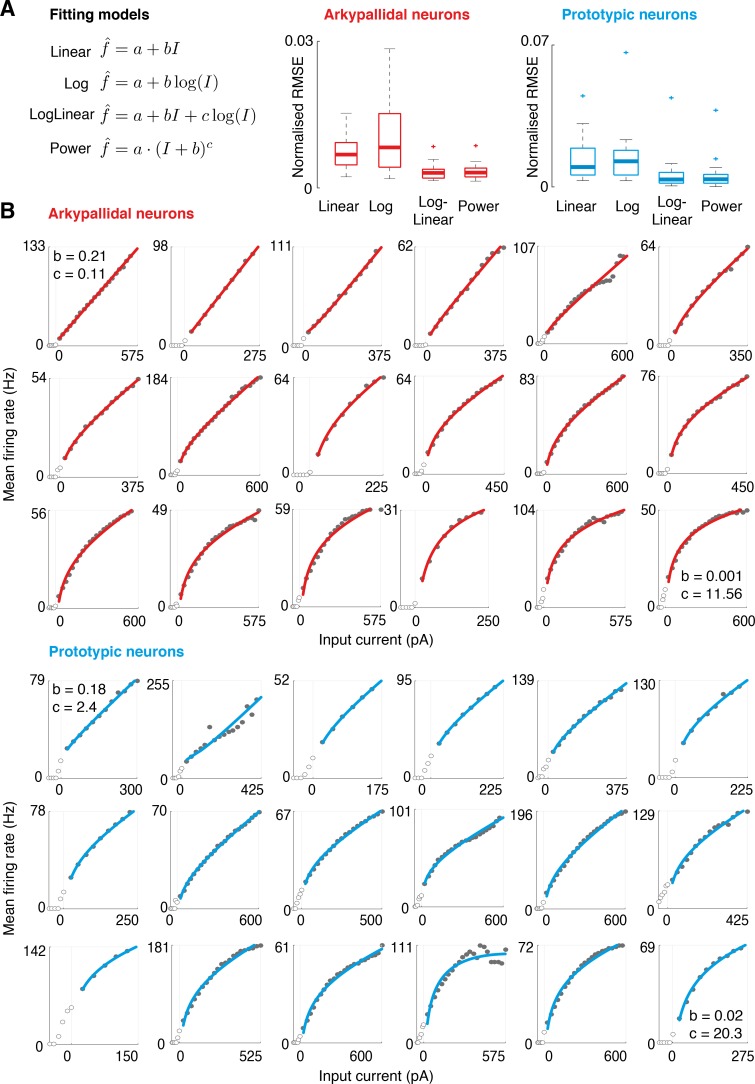
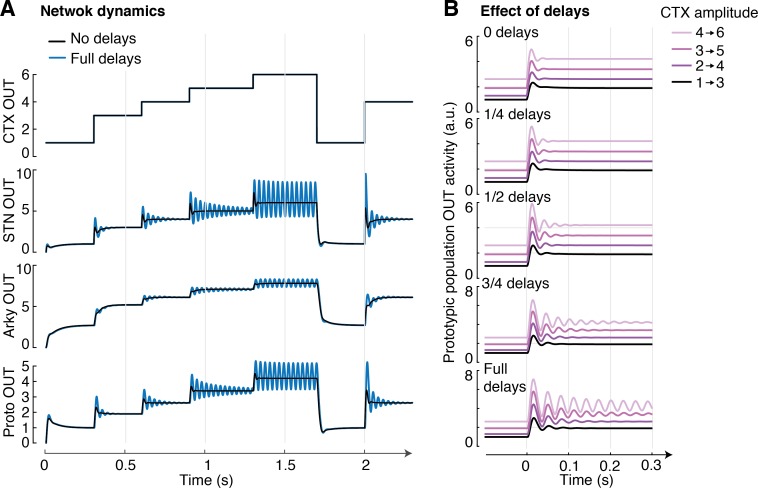
The external globus pallidus (GPe) is a key nucleus within basal ganglia circuits that are thought to be involved in action selection. A class of computational models assumes that, during action selection, the basal ganglia compute for all actions available in a given context the probabilities that they should be selected. These models suggest that a network of GPe and subthalamic nucleus (STN) neurons computes the normalization term in Bayes' equation. In order to perform such computation, the GPe needs to send feedback to the STN equal to a particular function of the activity of STN neurons. However, the complex form of this function makes it unlikely that individual GPe neurons, or even a single GPe cell type, could compute it. Here, we demonstrate how this function could be computed within a network containing two types of GABAergic GPe projection neuron, so-called 'prototypic' and 'arkypallidal' neurons, that have different response properties in vivo and distinct connections. We compare our model predictions with the experimentally-reported connectivity and input-output functions (f-I curves) of the two populations of GPe neurons. We show that, together, these dichotomous cell types fulfil the requirements necessary to compute the function needed for optimal action selection. We conclude that, by virtue of their distinct response properties and connectivities, a network of arkypallidal and prototypic GPe neurons comprises a neural substrate capable of supporting the computation of the posterior probabilities of actions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures






References
-
- Redgrave P, Prescott TJ, Gurney K. The basal ganglia: a vertebrate solution to the selection problem? Neuroscience 1999; 89: 1009–1023. - PubMed
-
- Kita H. Globus pallidus external segment. Prog Brain Res 2007; 160: 111–133. - PubMed
-
- Smith Y, Bevan MD, Shink E, Bolam JP. Microcircuitry of the direct and indirect pathways of the basal ganglia. Neuroscience 1998; 86: 353–387. - PubMed
-
- Frank MJ, Seeberger LC, O'Reilly R C. By carrot or by stick: cognitive reinforcement learning in parkinsonism. Science 2004; 306: 1940–1943. - PubMed
-
- Gurney K, Prescott TJ, Redgrave P. A computational model of action selection in the basal ganglia. I. A new functional anatomy. Biol Cybern 2001; 84: 401–410. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
