

HnRNP Q Has a Suppressive Role in the Translation of Mouse Cryptochrome1
- PMID: 27392095
- PMCID: PMC4938544
- DOI: 10.1371/journal.pone.0159018
HnRNP Q Has a Suppressive Role in the Translation of Mouse Cryptochrome1
Abstract
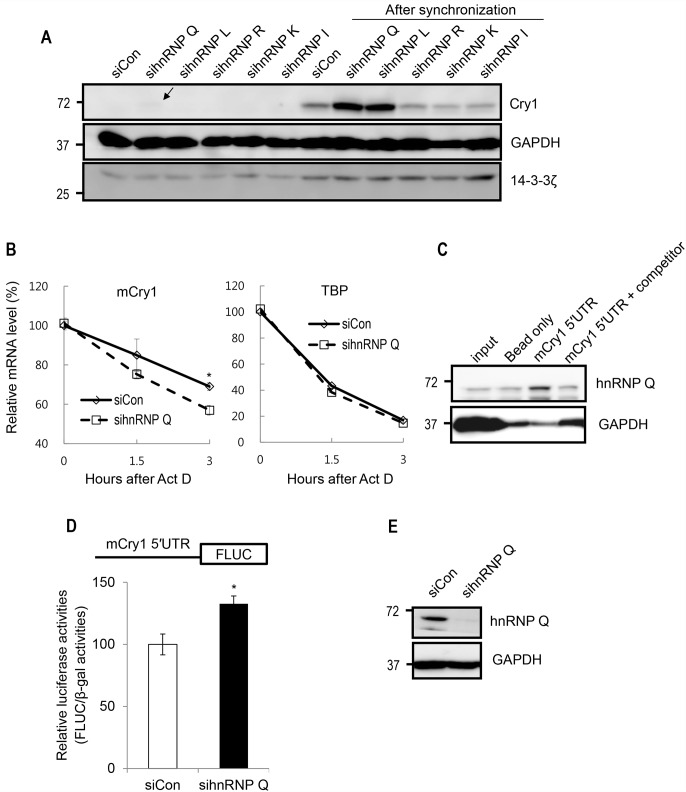
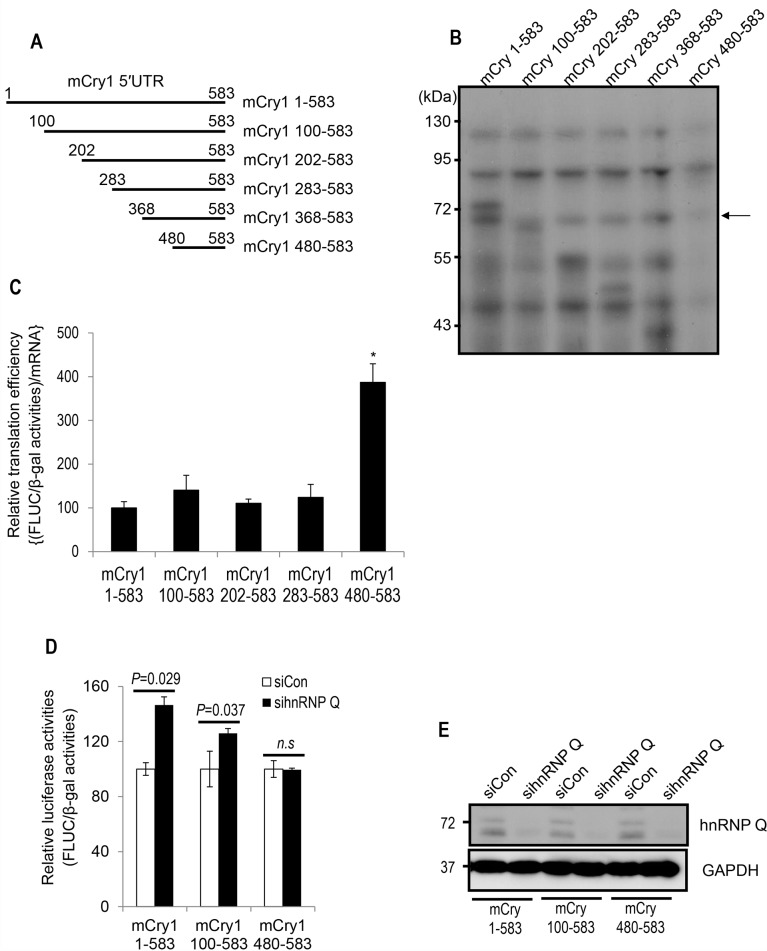
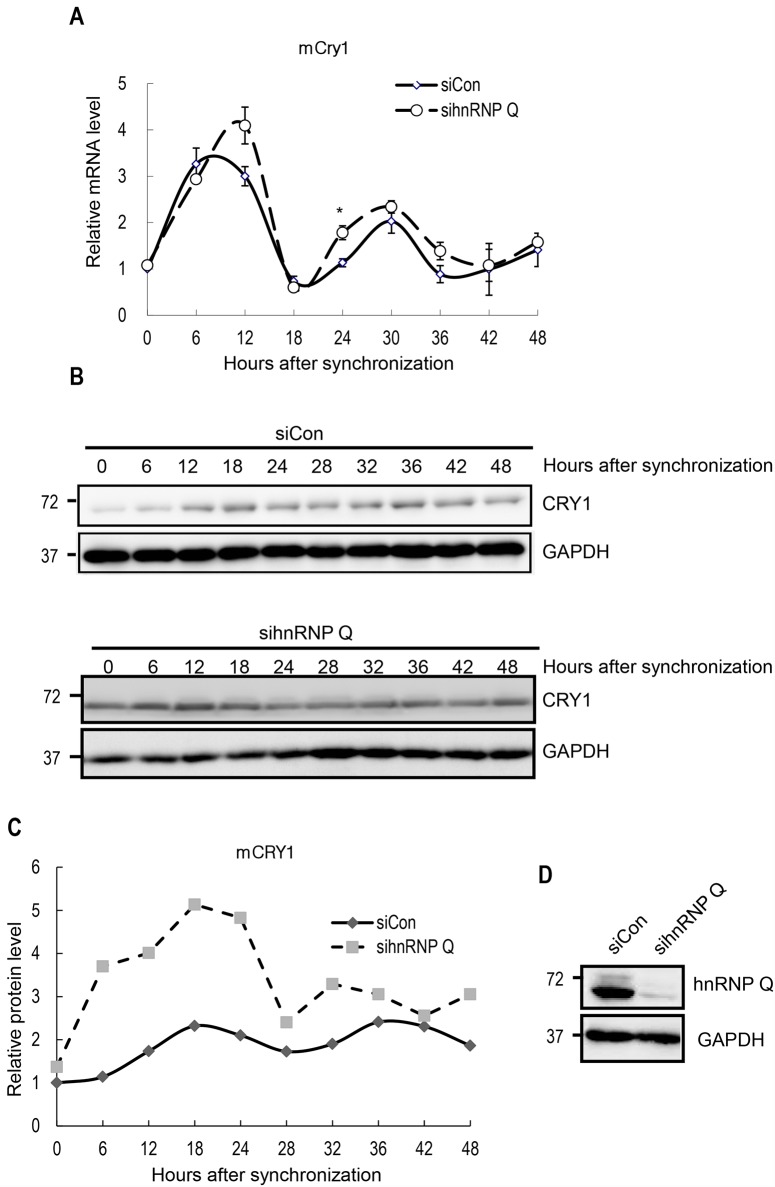
Precise regulation of gene expression is especially important for circadian timekeeping which is maintained by the proper oscillation of the mRNA and protein of clock genes and clock-controlled genes. As a main component of the core negative arm feedback loops in the circadian clock, the Cry1 gene contributes to the maintenance of behavioral and molecular rhythmicity. Despite the central role of Cry1, the molecular mechanisms regulating expression levels of Cry1 mRNA and protein are not well defined. In particular, the post-transcriptional regulation of Cry1 mRNA fate decisions is unclear. Here, we demonstrate that hnRNP Q binds to mCry1 mRNA via the 5'UTR. Furthermore, hnRNP Q inhibits the translation of mCry1 mRNA, leading to altered rhythmicity in the mCRY1 protein profile.
Conflict of interest statement
Figures
References
-
- Panda S, Hogenesch JB, Kay SA. Circadian rhythms from flies to human. Nature. 2002; 417: 329–335. - PubMed
-
- Barinaga M. Circadian rhythms. Two feedback loops run mammalian clock. Science. 2000; 288: 943–944. - PubMed
-
- Jin X, Shearman LP, Weaver DR, Zylka MJ, de Vries GJ, Reppert SM. A molecular mechanism regulating rhythmic output from the suprachiasmatic circadian clock. Cell. 1999; 96: 57–68. - PubMed
-
- Shearman LP, Sriram S, Weaver DR, Maywood ES, Chaves I, Zheng B, et al. Interacting molecular loops in the mammalian circadian clock. Science. 2000; 288: 1013–1019. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
