

Influenza virus RNA polymerase: insights into the mechanisms of viral RNA synthesis
- PMID: 27396566
- PMCID: PMC4966622
- DOI: 10.1038/nrmicro.2016.87
Influenza virus RNA polymerase: insights into the mechanisms of viral RNA synthesis
Abstract
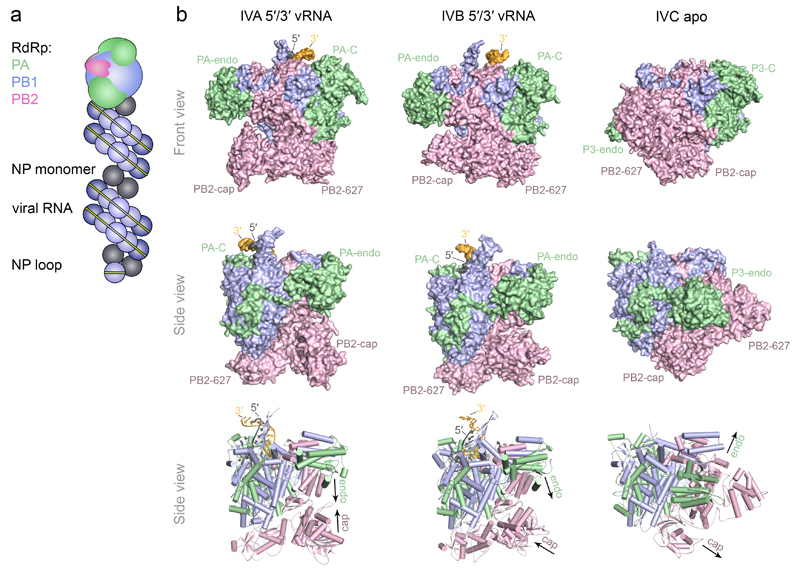
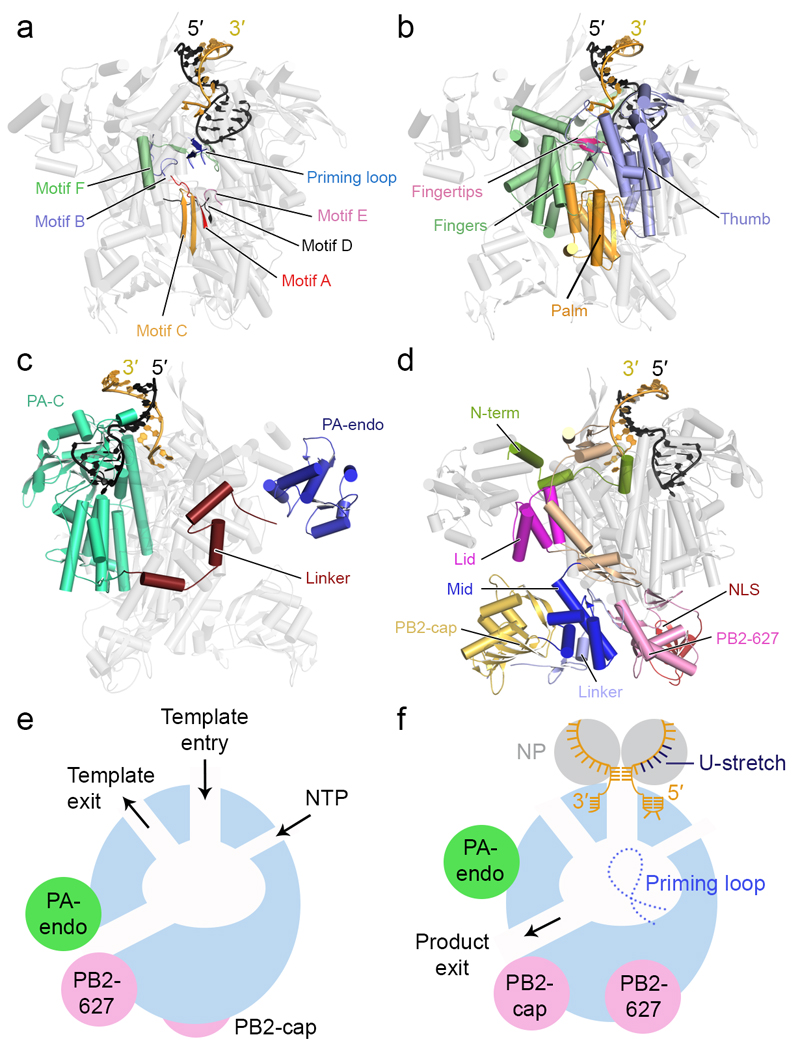
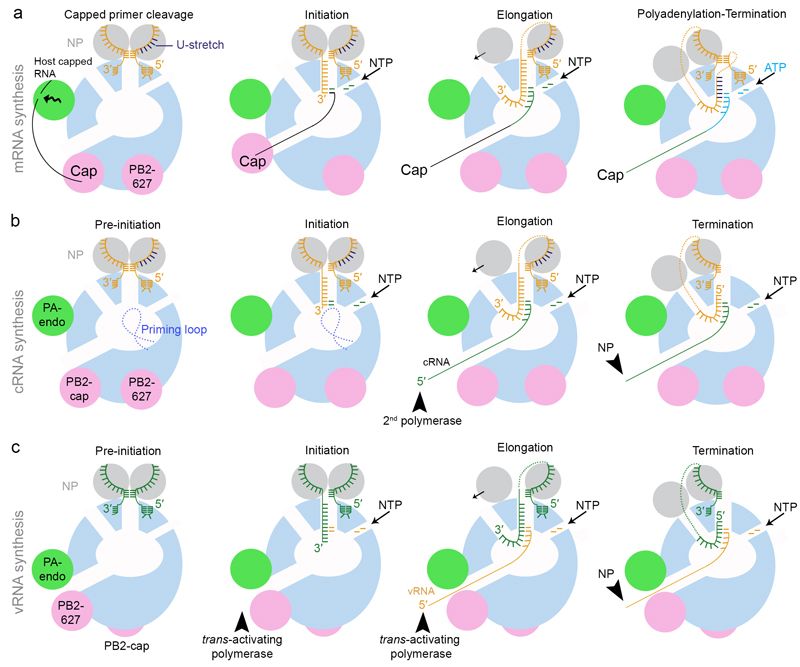
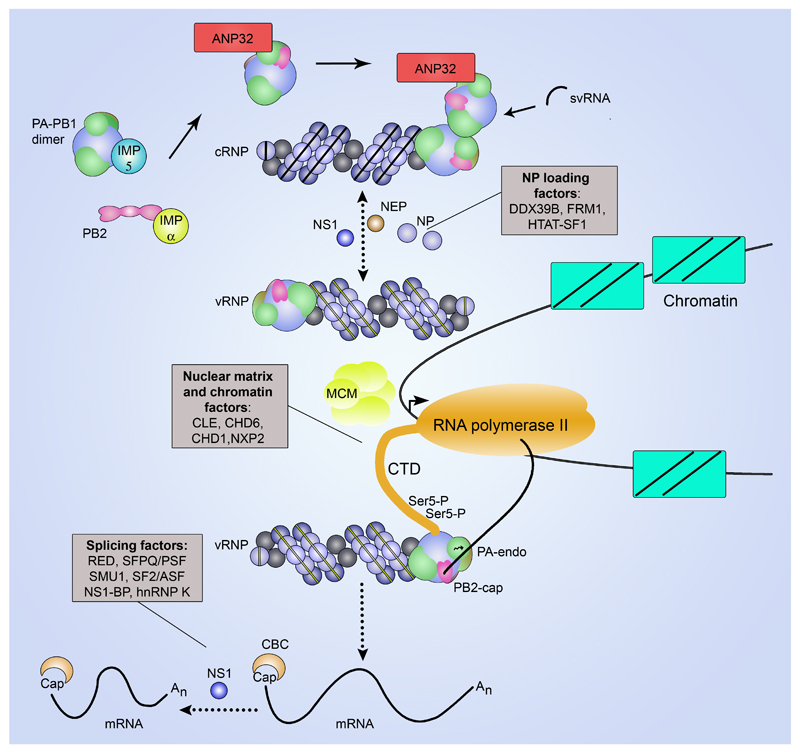
The genomes of influenza viruses consist of multiple segments of single-stranded negative-sense RNA. Each of these segments is bound by the heterotrimeric viral RNA-dependent RNA polymerase and multiple copies of nucleoprotein, which form viral ribonucleoprotein (vRNP) complexes. It is in the context of these vRNPs that the viral RNA polymerase carries out transcription of viral genes and replication of the viral RNA genome. In this Review, we discuss our current knowledge of the structure of the influenza virus RNA polymerase, and insights that have been gained into the molecular mechanisms of viral transcription and replication, and their regulation by viral and host factors. Furthermore, we discuss how advances in our understanding of the structure and function of polymerases could help in identifying new antiviral targets.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Molinari NA, et al. The annual impact of seasonal influenza in the US: measuring disease burden and costs. Vaccine. 2007;25:5086–96. - PubMed
-
- Muraki Y, Hongo S. The molecular virology and reverse genetics of influenza C virus. Jpn J Infect Dis. 2010;63:157–65. - PubMed
-
- Krammer F, Palese P. Advances in the development of influenza virus vaccines. Nat Rev Drug Discov. 2015;14:167–82. - PubMed
-
- Das K. Antivirals targeting influenza A virus. J Med Chem. 2012;55:6263–77. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
