

Distinct Functions of Argonaute Slicer in siRNA Maturation and Heterochromatin Formation
- PMID: 27397687
- PMCID: PMC5576859
- DOI: 10.1016/j.molcel.2016.05.039
Distinct Functions of Argonaute Slicer in siRNA Maturation and Heterochromatin Formation
Abstract
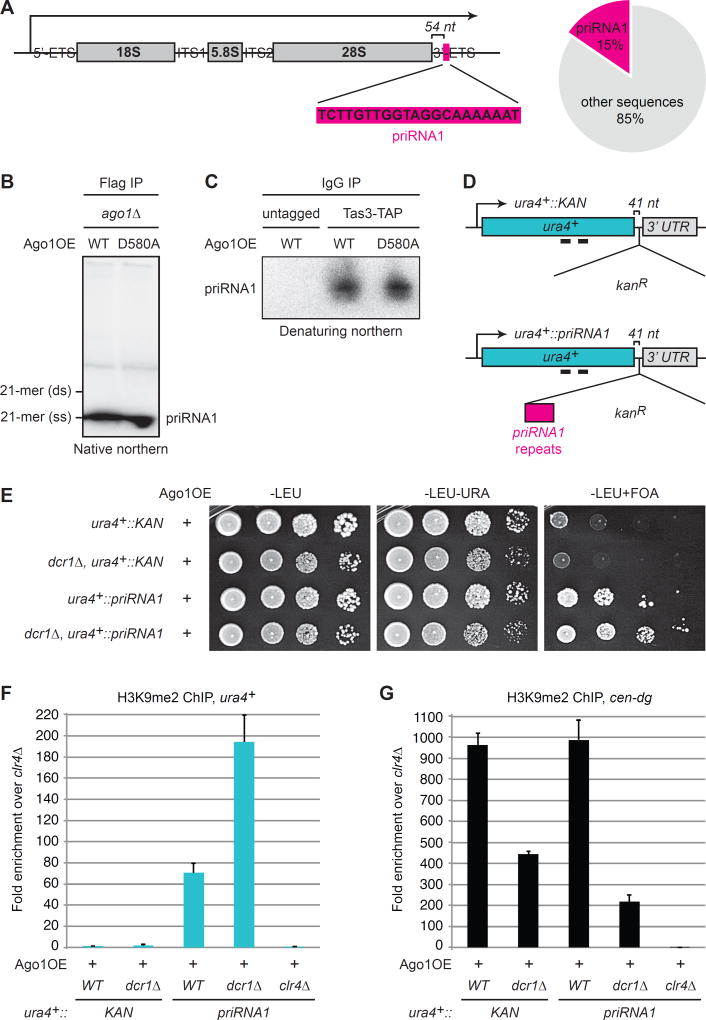
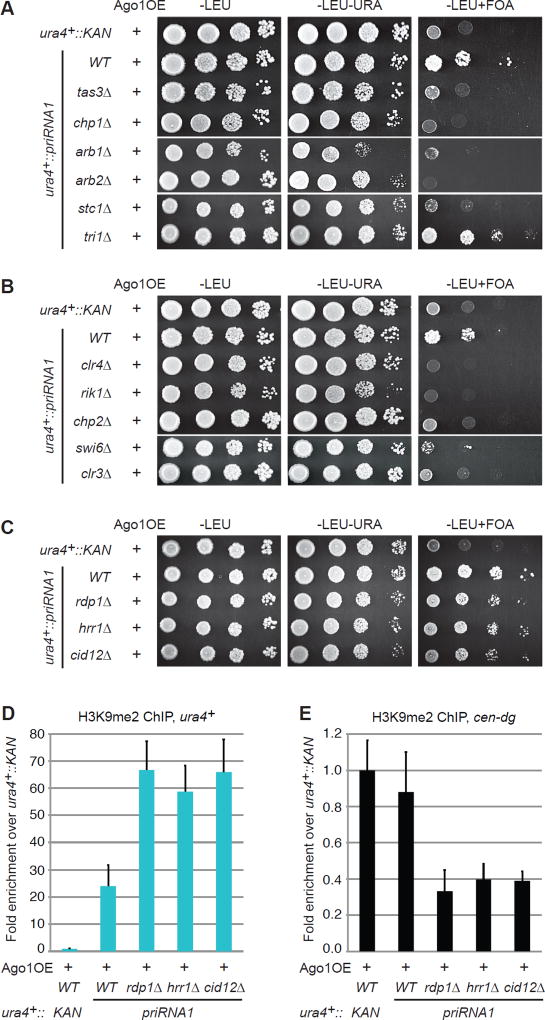
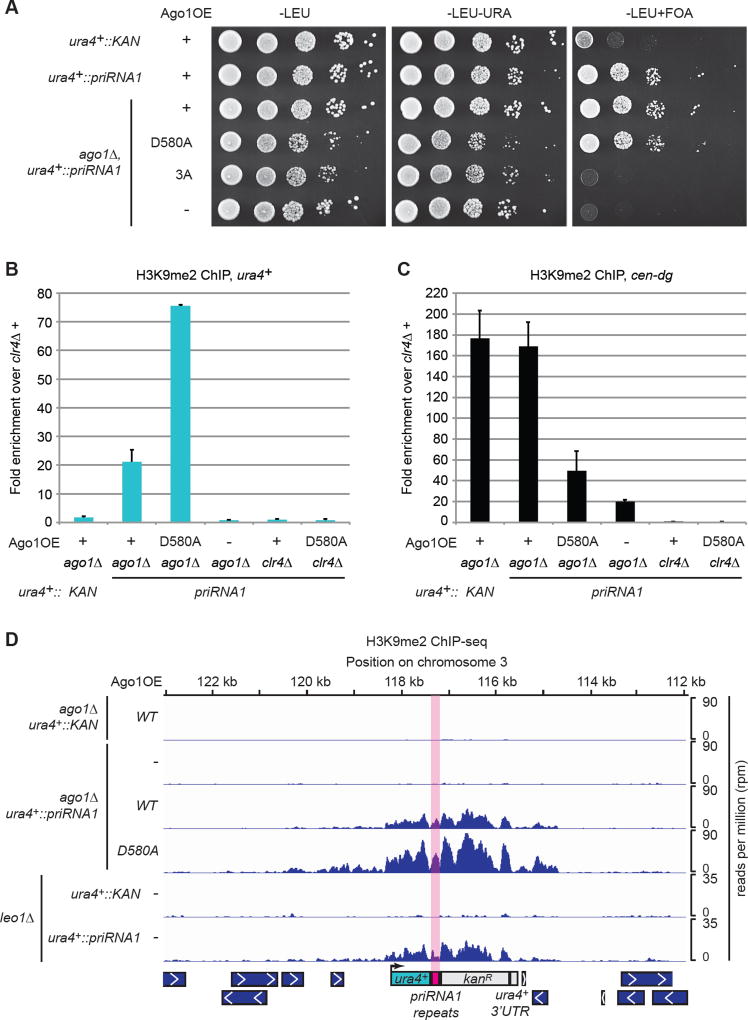
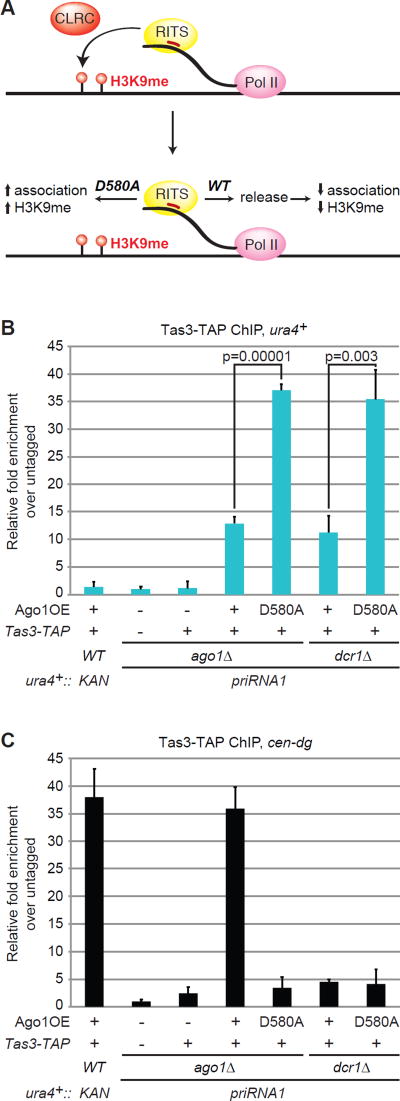
Small-RNA (sRNA)-guided transcriptional gene silencing by Argonaute (Ago)-containing complexes is fundamental to genome integrity and epigenetic inheritance. The RNA cleavage ("Slicer") activity of Argonaute has been implicated in both sRNA maturation and target RNA cleavage. Typically, Argonaute slices and releases the passenger strand of duplex sRNA to generate active silencing complexes, but it remains unclear whether slicing of target nascent RNAs, or other RNAi components, also contributes to downstream transcriptional silencing. Here, we develop a strategy for loading the fission yeast Ago1 with a single-stranded sRNA guide, which bypasses the requirement for slicer activity in generation of active silencing complexes. We show that slicer-defective Ago1 can mediate secondary sRNA generation, H3K9 methylation, and silencing similar to or better than wild-type and associates with chromatin more efficiently. The results define an ancient and minimal sRNA-mediated chromatin silencing mechanism, which resembles the germline-specific sRNA-dependent transcriptional silencing pathways in Drosophila and mammals.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures



References
-
- Aravin AA, Hannon GJ, Brennecke J. The Piwi-piRNA pathway provides an adaptive defense in the transposon arms race. Science. 2007;318:761–764. - PubMed
-
- Bahler J, Wu JQ, Longtine MS, Shah NG, McKenzie A, 3rd, Steever AB, Wach A, Philippsen P, Pringle JR. Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast. 1998;14:943–951. - PubMed
-
- Bannister AJ, Zegerman P, Partridge JF, Miska EA, Thomas JO, Allshire RC, Kouzarides T. Selective recognition of methylated lysine 9 on histone H3 by the HP1 chromo domain. Nature. 2001;410:120–124. - PubMed
-
- Bartel DP. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell. 2004;116:281–297. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
