

Redox Nanodomains Are Induced by and Control Calcium Signaling at the ER-Mitochondrial Interface
- PMID: 27397688
- PMCID: PMC4998968
- DOI: 10.1016/j.molcel.2016.05.040
Redox Nanodomains Are Induced by and Control Calcium Signaling at the ER-Mitochondrial Interface
Abstract
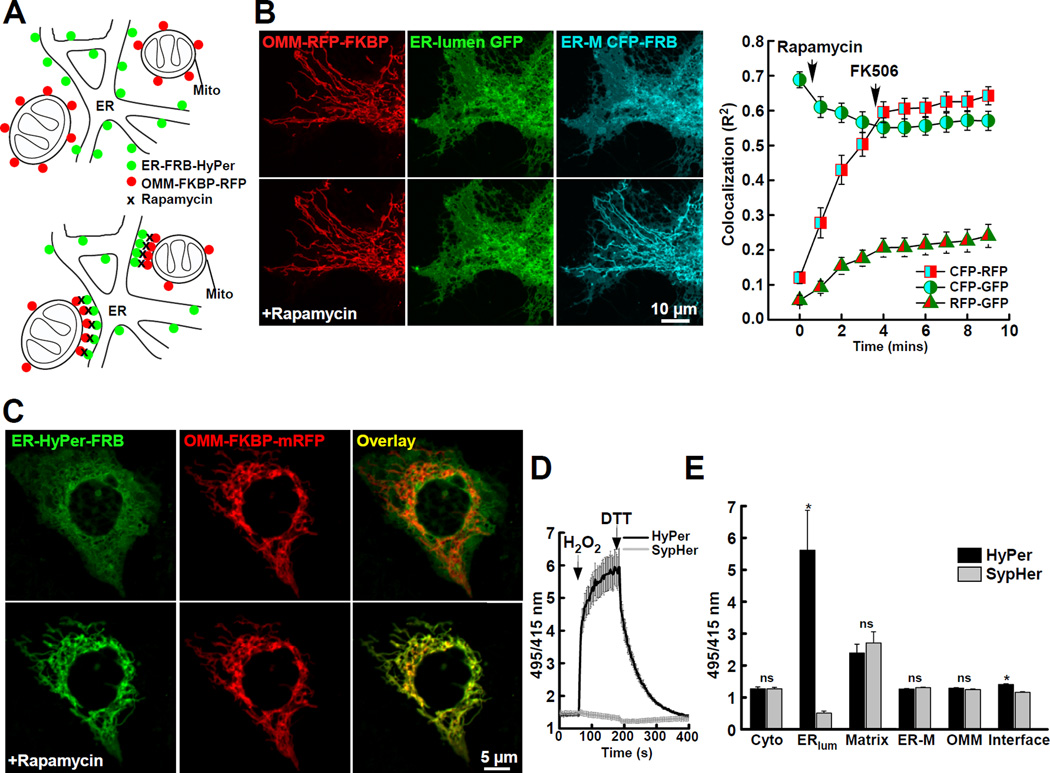
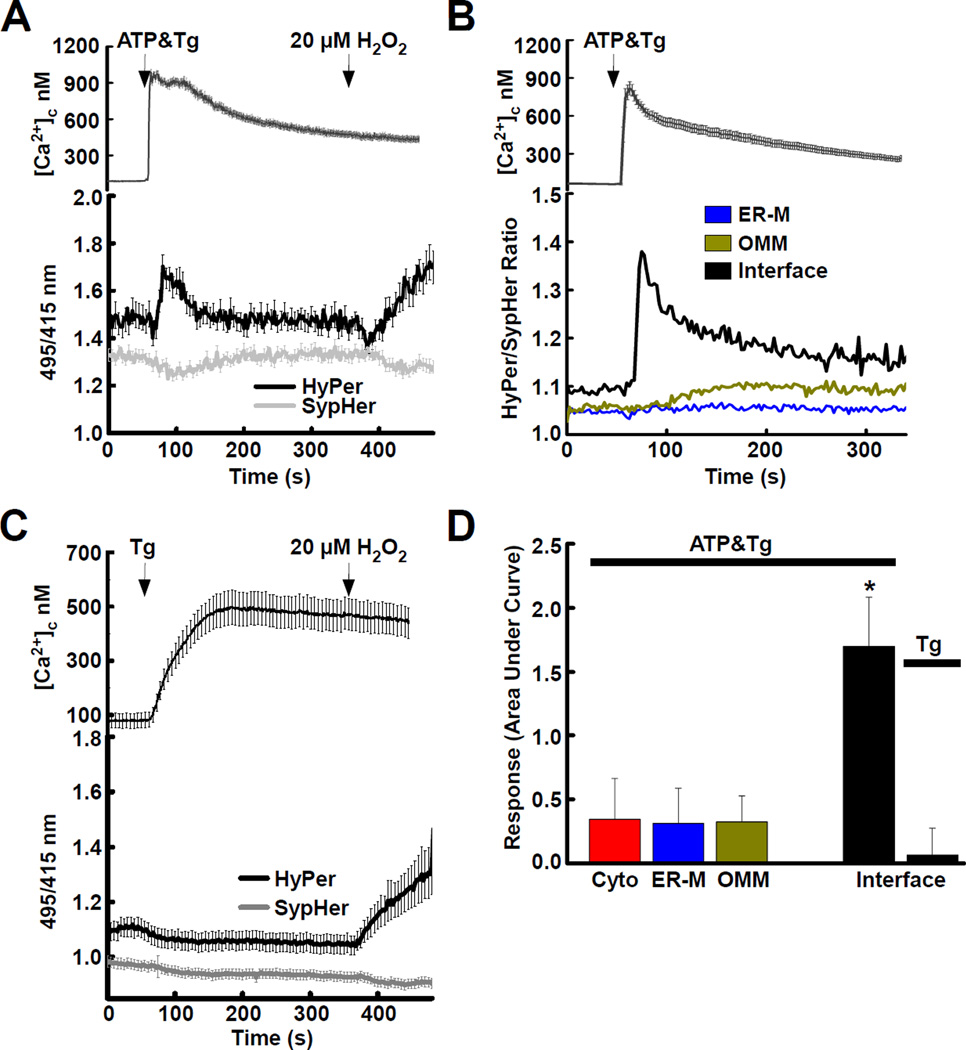
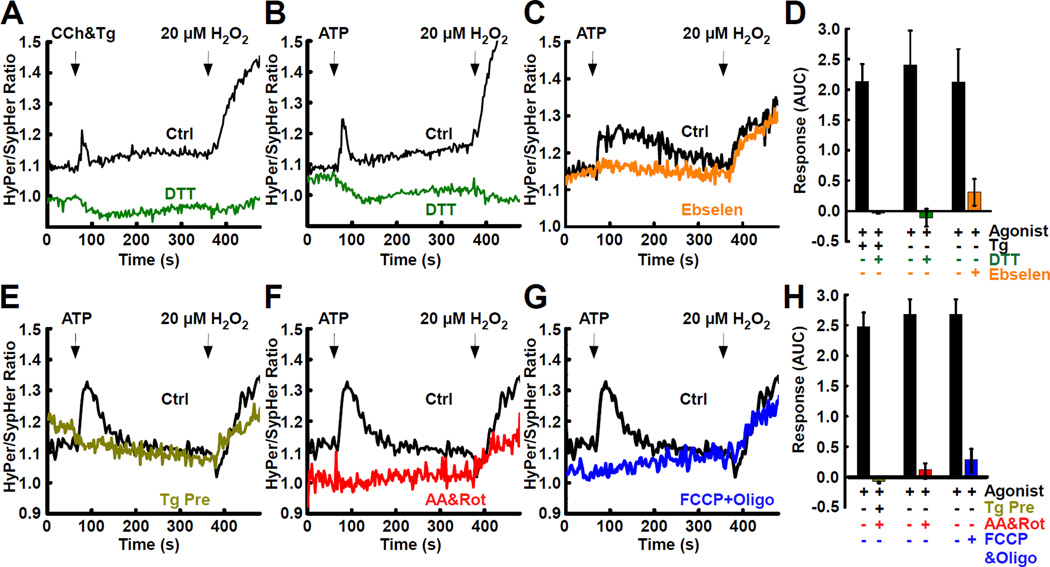
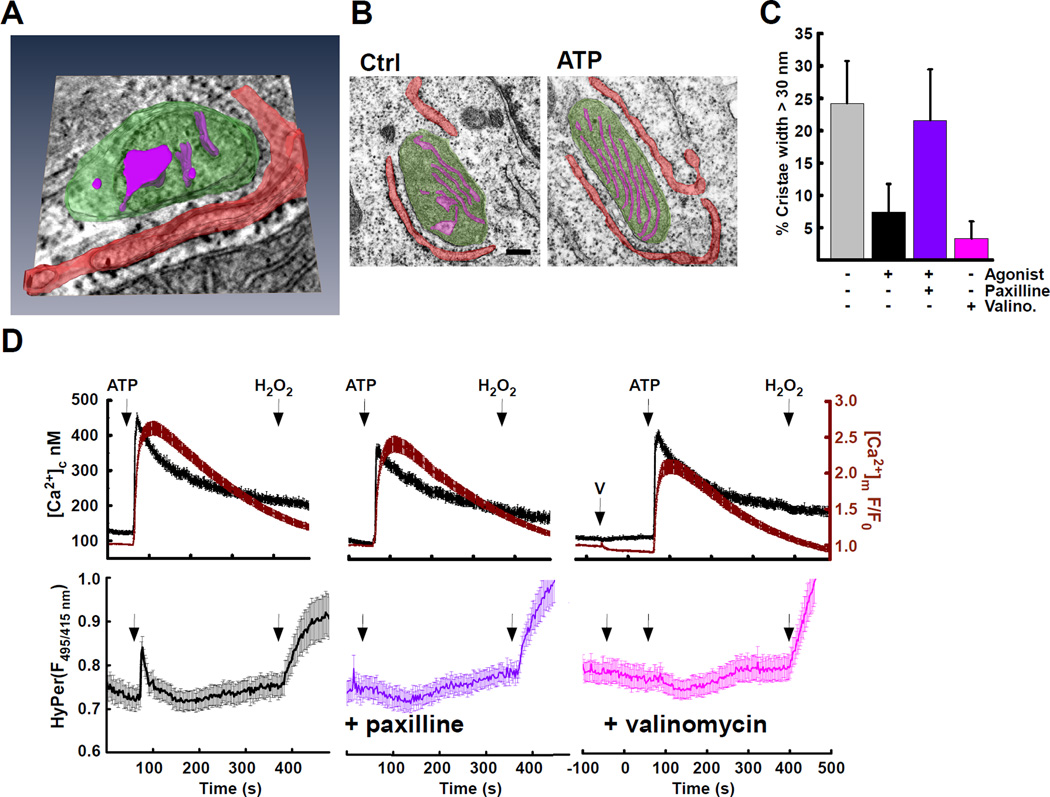
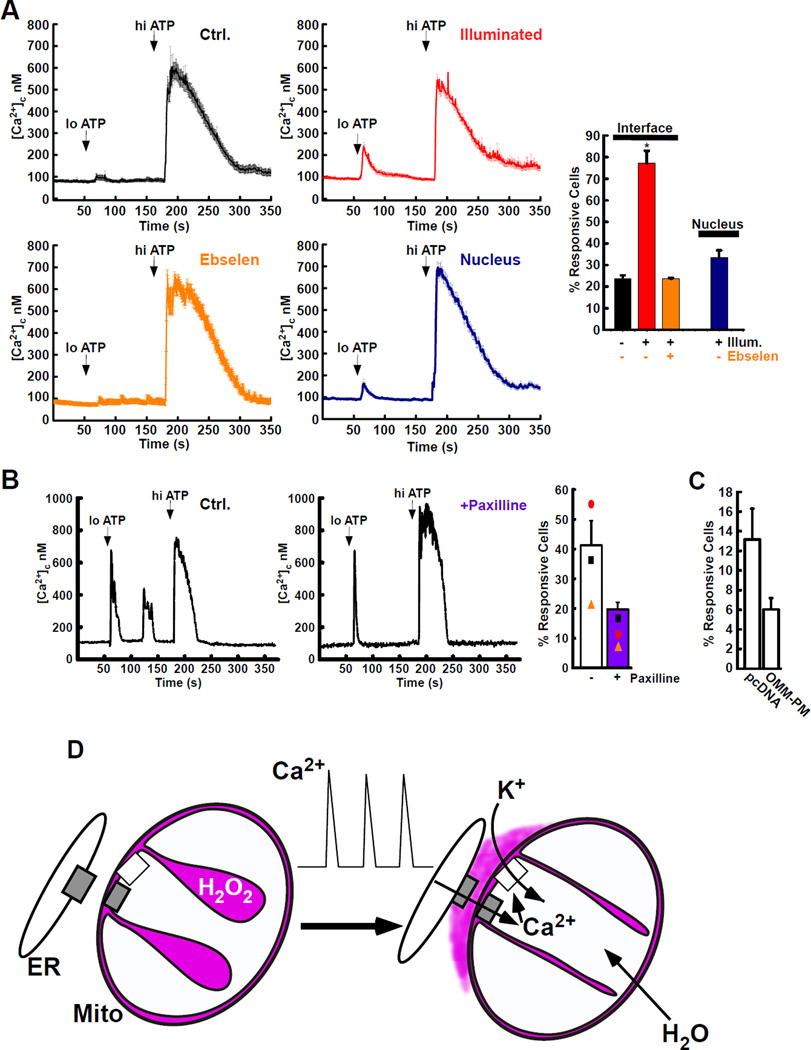
The ER-mitochondrial interface is central to calcium signaling, organellar dynamics, and lipid biosynthesis. The ER and mitochondrial membranes also host sources and targets of reactive oxygen species (ROS), but their local dynamics and relevance remained elusive since measurement and perturbation of ROS at the organellar interface has proven difficult. Employing drug-inducible synthetic ER-mitochondrial linkers, we overcame this problem and demonstrate that the ER-mitochondrial interface hosts a nanodomain of H2O2, which is induced by cytoplasmic [Ca(2+)] spikes and exerts a positive feedback on calcium oscillations. H2O2 nanodomains originate from the mitochondrial cristae, which are compressed upon calcium signal propagation to the mitochondria, likely due to Ca(2+)-induced K(+) and concomitant water influx to the matrix. Thus, ER-mitochondrial H2O2 nanodomains represent a component of inter-organelle communication, regulating calcium signaling and mitochondrial activities.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Calcium and redox signals at mitochondrial interfaces: A nanoview perspective.Cell Calcium. 2022 May;103:102550. doi: 10.1016/j.ceca.2022.102550. Epub 2022 Feb 4. Cell Calcium. 2022. PMID: 35144095 No abstract available.
References
-
- Aon MA, Cortassa S, Marban E, O'Rourke B. Synchronized whole cell oscillations in mitochondrial metabolism triggered by a local release of reactive oxygen species in cardiac myocytes. The Journal of biological chemistry. 2003;278:44735–44744. - PubMed
-
- Bader M, Muse W, Ballou DP, Gassner C, Bardwell JC. Oxidative protein folding is driven by the electron transport system. Cell. 1999;98:217–227. - PubMed
-
- Belousov VV, Fradkov AF, Lukyanov KA, Staroverov DB, Shakhbazov KS, Terskikh AV, Lukyanov S. Genetically encoded fluorescent indicator for intracellular hydrogen peroxide. Nature methods. 2006;3:281–286. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
