

HCN-channel dendritic targeting requires bipartite interaction with TRIP8b and regulates antidepressant-like behavioral effects
- PMID: 27400855
- PMCID: PMC5233690
- DOI: 10.1038/mp.2016.99
HCN-channel dendritic targeting requires bipartite interaction with TRIP8b and regulates antidepressant-like behavioral effects
Abstract
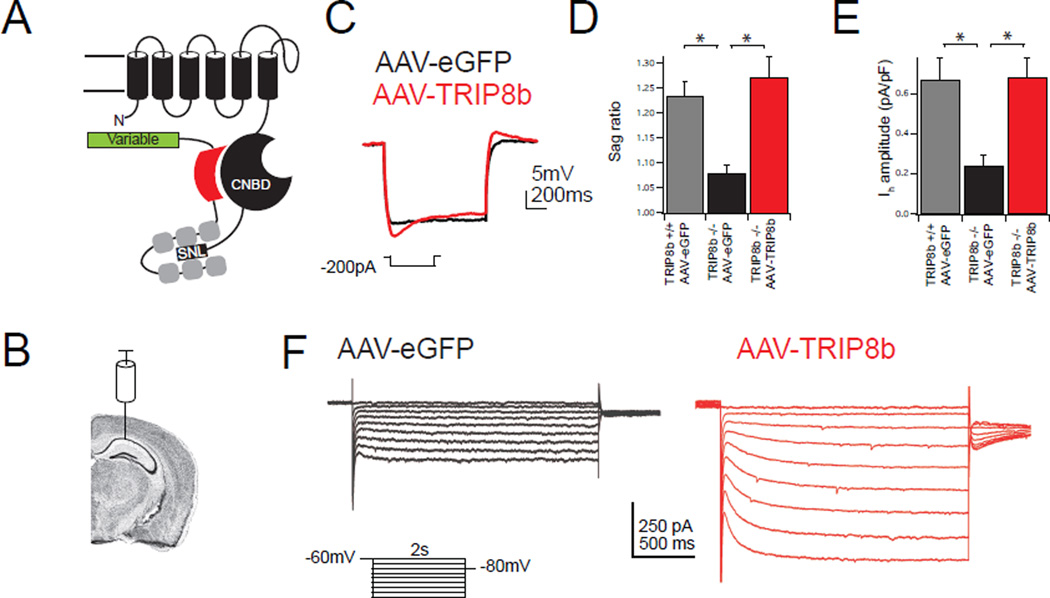
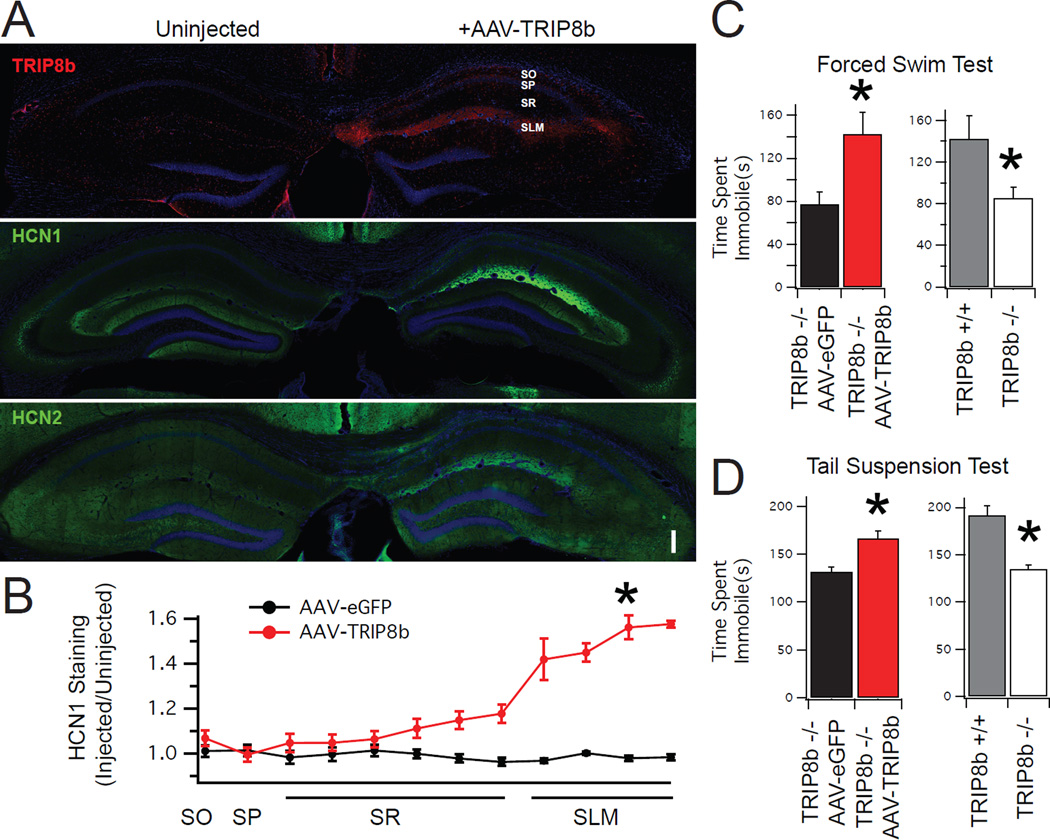
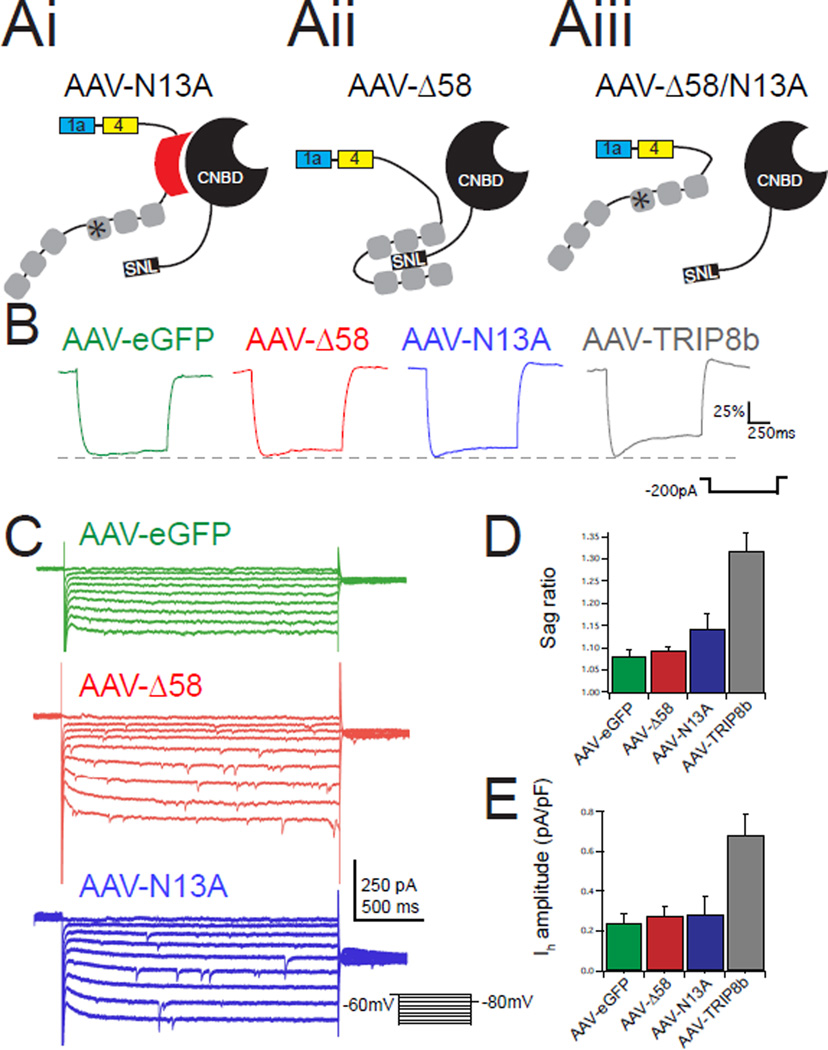
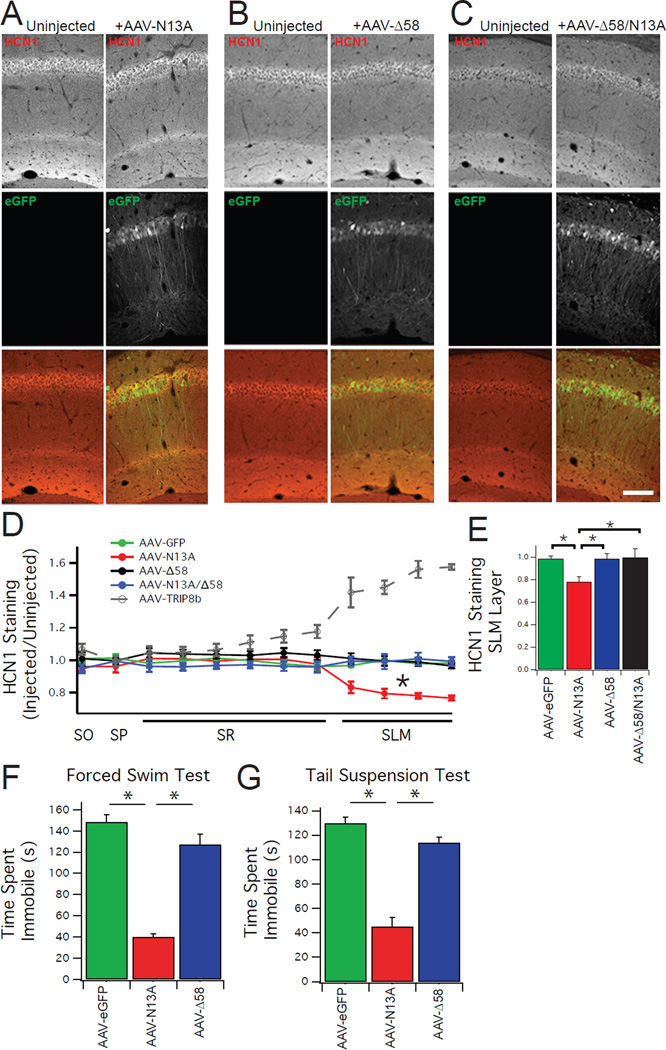
Major depressive disorder (MDD) is a prevalent psychiatric condition with limited therapeutic options beyond monoaminergic therapies. Although effective in some individuals, many patients fail to respond adequately to existing treatments, and new pharmacologic targets are needed. Hyperpolarization-activated cyclic nucleotide-gated (HCN) channels regulate excitability in neurons, and blocking HCN channel function has been proposed as a novel antidepressant strategy. However, systemic blockade of HCN channels produces cardiac effects that limit this approach. Knockout (KO) of the brain-specific HCN-channel auxiliary subunit tetratricopeptide repeat-containing Rab8b-interacting protein (TRIP8b) also produces antidepressant-like behavioral effects and suggests that inhibiting TRIP8b function could produce antidepressant-like effects without affecting the heart. We examined the structural basis of TRIP8b-mediated HCN-channel trafficking and its relationship with antidepressant-like behavior using a viral rescue approach in TRIP8b KO mice. We found that restoring TRIP8b to the hippocampus was sufficient to reverse the impaired HCN-channel trafficking and antidepressant-like behavioral effects caused by TRIP8b KO. Moreover, we found that hippocampal expression of a mutated version of TRIP8b further impaired HCN-channel trafficking and increased the antidepressant-like behavioral phenotype of TRIP8b KO mice. Thus, modulating the TRIP8b-HCN interaction bidirectionally influences channel trafficking and antidepressant-like behavior. Overall, our work suggests that small-molecule inhibitors of the interaction between TRIP8b and HCN should produce antidepressant-like behaviors and could represent a new paradigm for the treatment of MDD.
Conflict of interest statement
Conflict of Interests Statement The authors declare no conflicts of interest.
Figures
Similar articles
-
Deletion of the hyperpolarization-activated cyclic nucleotide-gated channel auxiliary subunit TRIP8b impairs hippocampal Ih localization and function and promotes antidepressant behavior in mice.J Neurosci. 2011 May 18;31(20):7424-40. doi: 10.1523/JNEUROSCI.0936-11.2011. J Neurosci. 2011. PMID: 21593326 Free PMC article.
-
Trafficking and gating of hyperpolarization-activated cyclic nucleotide-gated channels are regulated by interaction with tetratricopeptide repeat-containing Rab8b-interacting protein (TRIP8b) and cyclic AMP at distinct sites.J Biol Chem. 2011 Jun 10;286(23):20823-34. doi: 10.1074/jbc.M111.236125. Epub 2011 Apr 19. J Biol Chem. 2011. PMID: 21504900 Free PMC article.
-
Method for Identifying Small Molecule Inhibitors of the Protein-protein Interaction Between HCN1 and TRIP8b.J Vis Exp. 2016 Nov 11;(117):54540. doi: 10.3791/54540. J Vis Exp. 2016. PMID: 27911380 Free PMC article.
-
HCN channels: New targets for the design of an antidepressant with rapid effects.J Affect Disord. 2019 Feb 15;245:764-770. doi: 10.1016/j.jad.2018.11.081. Epub 2018 Nov 13. J Affect Disord. 2019. PMID: 30448761 Review.
-
The structure and function of TRIP8b, an auxiliary subunit of hyperpolarization-activated cyclic-nucleotide gated channels.Channels (Austin). 2020 Dec;14(1):110-122. doi: 10.1080/19336950.2020.1740501. Channels (Austin). 2020. PMID: 32189562 Free PMC article. Review.
Cited by
-
Perisomatic changes in h-channels regulate depressive behaviors following chronic unpredictable stress.Mol Psychiatry. 2018 Apr;23(4):892-903. doi: 10.1038/mp.2017.28. Epub 2017 Apr 18. Mol Psychiatry. 2018. PMID: 28416809 Free PMC article.
-
Hippocampal dorsal CA1: Functional connectivity and role in HCN channelopathies in affective diseases and epilepsy.IBRO Neurosci Rep. 2025 Apr 4;18:644-656. doi: 10.1016/j.ibneur.2025.03.008. eCollection 2025 Jun. IBRO Neurosci Rep. 2025. PMID: 40292082 Free PMC article.
-
Regulation of HCN Channels by Protein Interactions.Front Physiol. 2022 Jun 20;13:928507. doi: 10.3389/fphys.2022.928507. eCollection 2022. Front Physiol. 2022. PMID: 35795651 Free PMC article. Review.
-
Discovery of a small-molecule inhibitor of the TRIP8b-HCN interaction with efficacy in neurons.J Biol Chem. 2022 Jul;298(7):102069. doi: 10.1016/j.jbc.2022.102069. Epub 2022 May 24. J Biol Chem. 2022. PMID: 35623388 Free PMC article.
-
Hyperpolarization-Activated Cyclic Nucleotide-Gated Channels: An Emerging Role in Neurodegenerative Diseases.Front Mol Neurosci. 2019 Jun 5;12:141. doi: 10.3389/fnmol.2019.00141. eCollection 2019. Front Mol Neurosci. 2019. PMID: 31231190 Free PMC article. Review.
References
-
- Kessler RC, Berglund P, Demler O, Jin R, Merikangas KR, Walters EE. Lifetime prevalence and age-of-onset distributions of DSM-IV disorders in the National Comorbidity Survey Replication. Arch Gen Psychiatry. 2005;62:593–602. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
