

Whole-Genome Sequencing of Native Sheep Provides Insights into Rapid Adaptations to Extreme Environments
- PMID: 27401233
- PMCID: PMC5026255
- DOI: 10.1093/molbev/msw129
Whole-Genome Sequencing of Native Sheep Provides Insights into Rapid Adaptations to Extreme Environments
Abstract
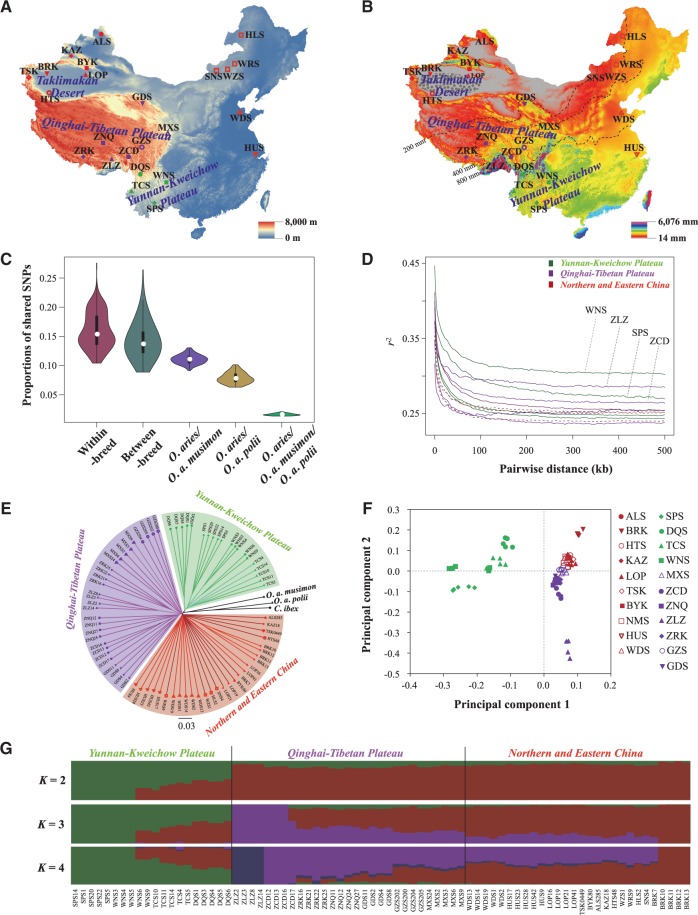
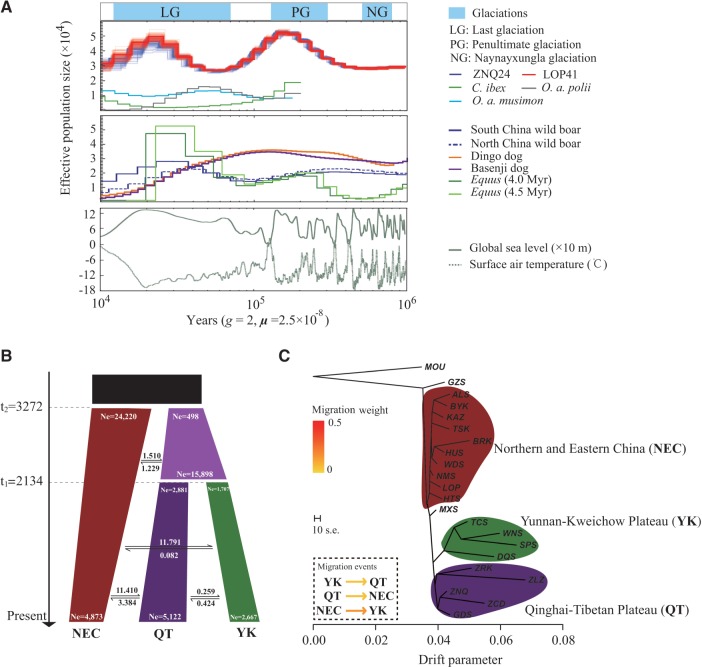
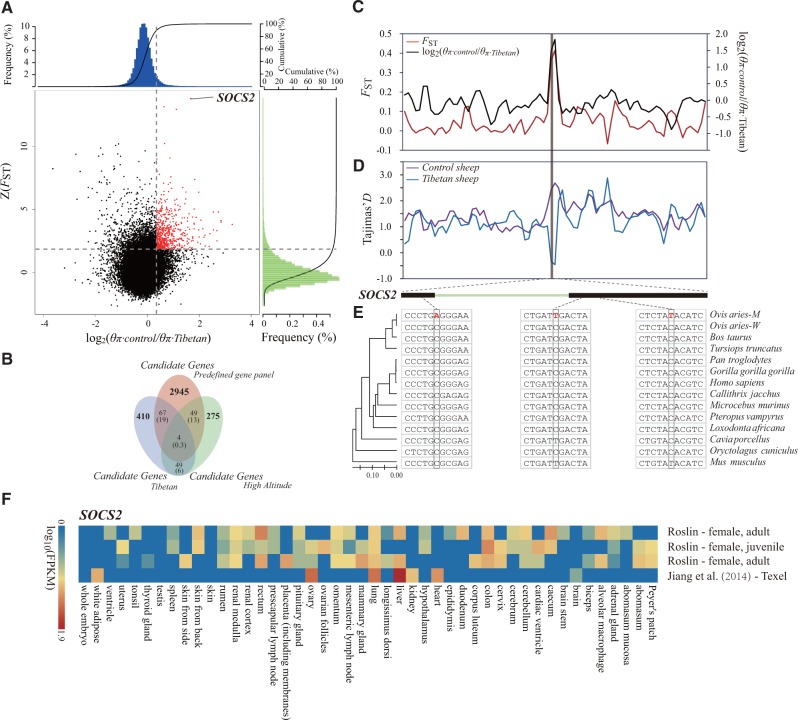
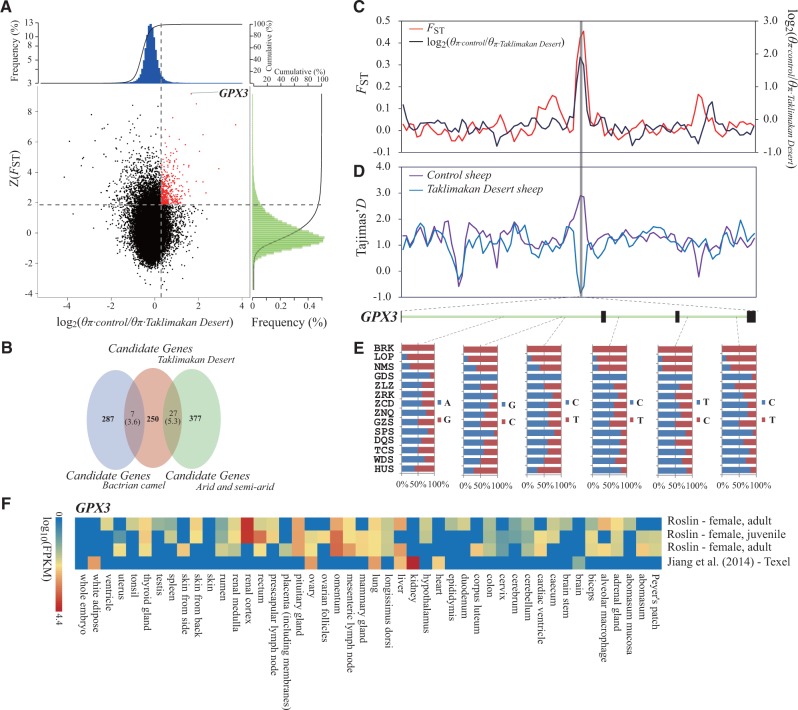
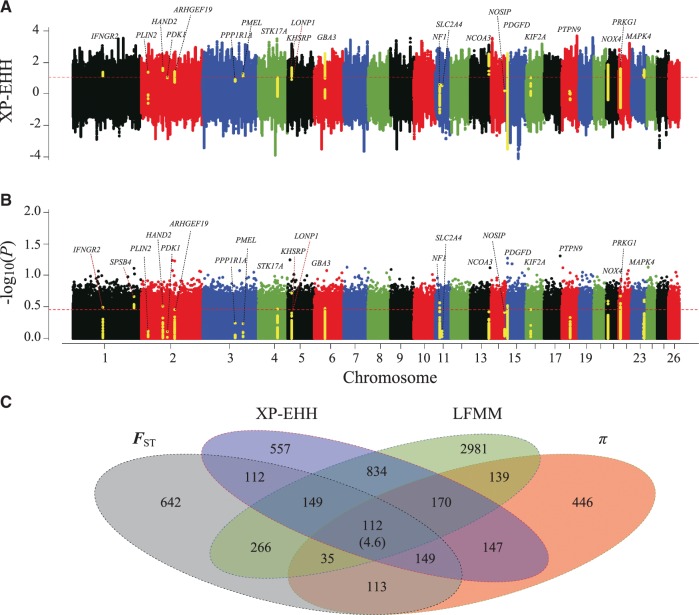
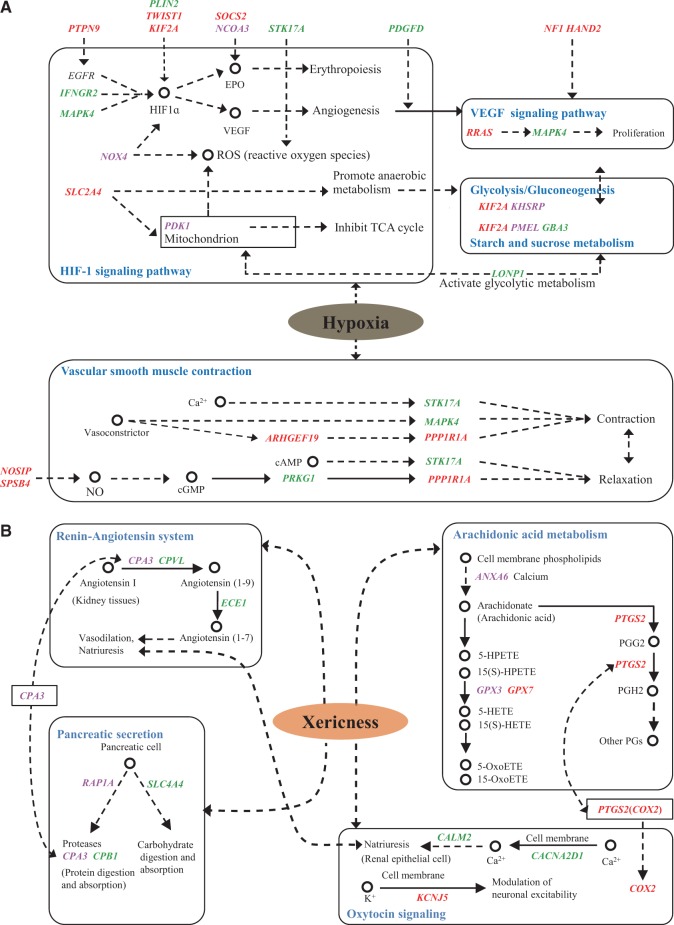
Global climate change has a significant effect on extreme environments and a profound influence on species survival. However, little is known of the genome-wide pattern of livestock adaptations to extreme environments over a short time frame following domestication. Sheep (Ovis aries) have become well adapted to a diverse range of agroecological zones, including certain extreme environments (e.g., plateaus and deserts), during their post-domestication (approximately 8-9 kya) migration and differentiation. Here, we generated whole-genome sequences from 77 native sheep, with an average effective sequencing depth of ∼5× for 75 samples and ∼42× for 2 samples. Comparative genomic analyses among sheep in contrasting environments, that is, plateau (>4,000 m above sea level) versus lowland (<100 m), high-altitude region (>1500 m) versus low-altitude region (<1300 m), desert (<10 mm average annual precipitation) versus highly humid region (>600 mm), and arid zone (<400 mm) versus humid zone (>400 mm), detected a novel set of candidate genes as well as pathways and GO categories that are putatively associated with hypoxia responses at high altitudes and water reabsorption in arid environments. In addition, candidate genes and GO terms functionally related to energy metabolism and body size variations were identified. This study offers novel insights into rapid genomic adaptations to extreme environments in sheep and other animals, and provides a valuable resource for future research on livestock breeding in response to climate change.
Keywords: Ovis aries; climate change.; extreme environment; rapid adaptation; whole-genome sequencing.
© The Author 2016. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures

References
-
- Barrett JC, Fry B, Maller J, Daly MJ. 2005. Haploview: analysis and visualization of LD and haplotype maps. Bioinformatics 21:263–265. - PubMed
-
- Breyer MD, Breyer RM. 2000. Prostaglandin receptors: their role in regulating renal function. Curr Opin Nephrol Hypertens. 9:23–29. - PubMed
-
- Brown ME, Funk CC. 2008. Food security under climate change. Science 319:580–581. - PubMed
-
- Carroll MA, Balazy M, Margiotta P, Huang DD, Falck JR, McGiff JC. 1996. Cytochrome P-450-dependent HETEs: profile of biological activity and stimulation by vasoactive peptides. Am J Physiol. 271:R863–R869. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
