

HGF-independent regulation of MET and GAB1 by nonreceptor tyrosine kinase FER potentiates metastasis in ovarian cancer
- PMID: 27401557
- PMCID: PMC4949327
- DOI: 10.1101/gad.284166.116
HGF-independent regulation of MET and GAB1 by nonreceptor tyrosine kinase FER potentiates metastasis in ovarian cancer
Abstract
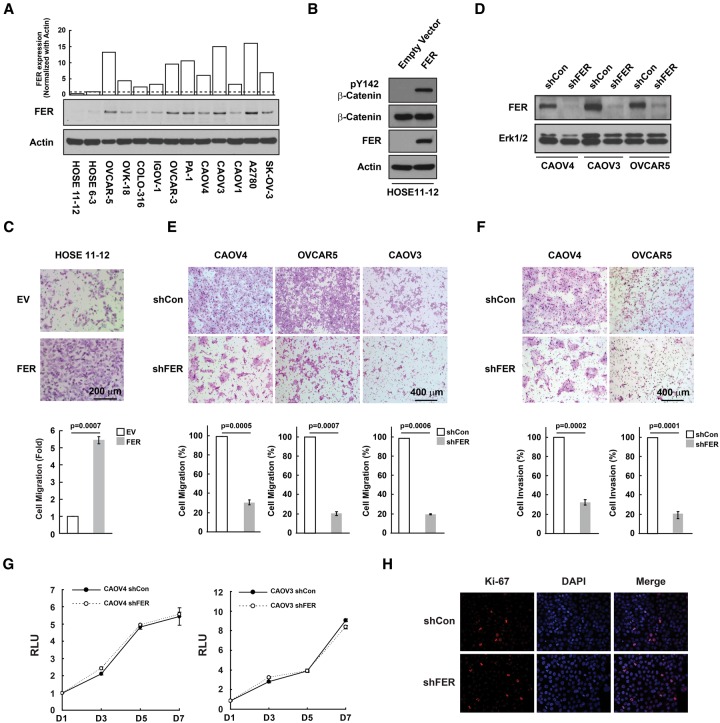
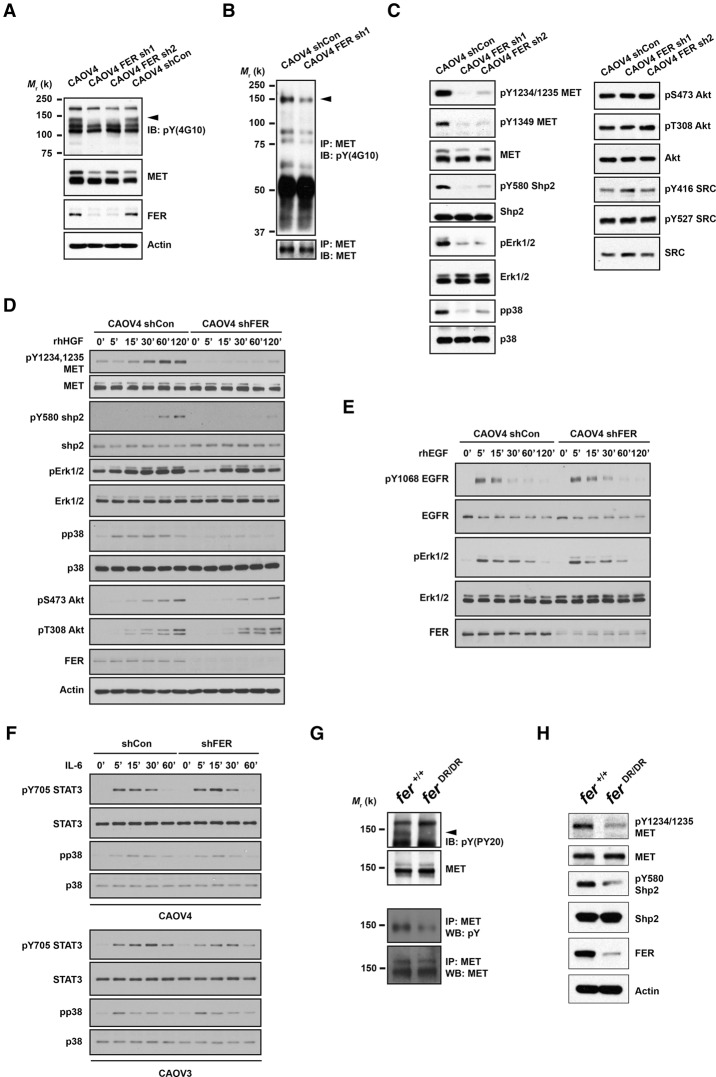
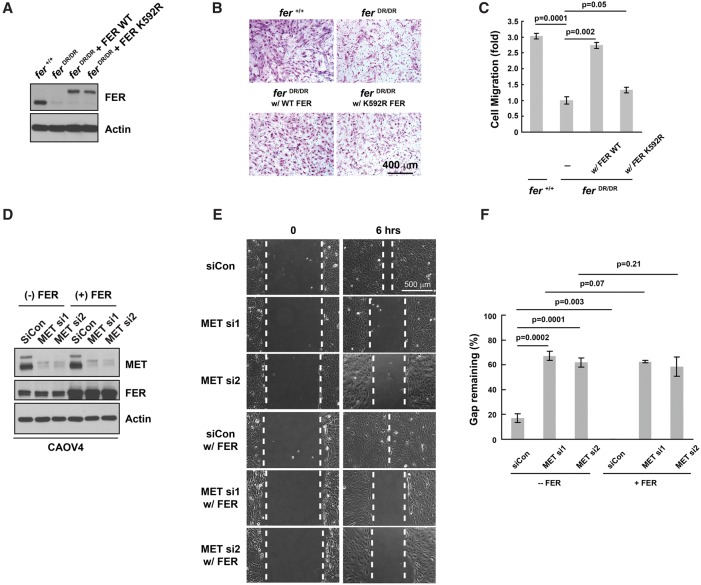
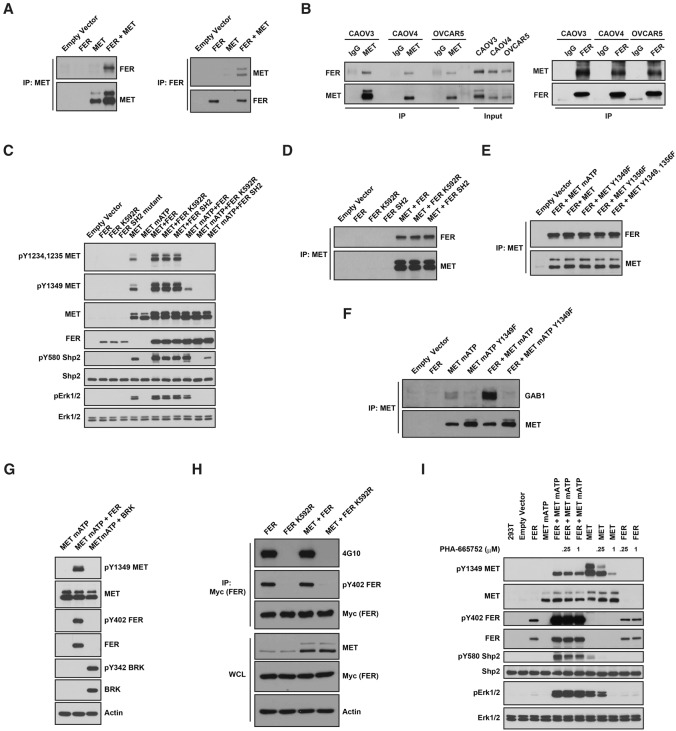
Ovarian cancer cells disseminate readily within the peritoneal cavity, which promotes metastasis, and are often resistant to chemotherapy. Ovarian cancer patients tend to present with advanced disease, which also limits treatment options; consequently, new therapies are required. The oncoprotein tyrosine kinase MET, which is the receptor for hepatocyte growth factor (HGF), has been implicated in ovarian tumorigenesis and has been the subject of extensive drug development efforts. Here, we report a novel ligand- and autophosphorylation-independent activation of MET through the nonreceptor tyrosine kinase feline sarcoma-related (FER). We demonstrated that the levels of FER were elevated in ovarian cancer cell lines relative to those in immortalized normal surface epithelial cells and that suppression of FER attenuated the motility and invasive properties of these cancer cells. Furthermore, loss of FER impaired the metastasis of ovarian cancer cells in vivo. Mechanistically, we demonstrated that FER phosphorylated a signaling site in MET: Tyr1349. This enhanced activation of RAC1/PAK1 and promoted a kinase-independent scaffolding function that led to recruitment and phosphorylation of GAB1 and the specific activation of the SHP2-ERK signaling pathway. Overall, this analysis provides new insights into signaling events that underlie metastasis in ovarian cancer cells, consistent with a prometastatic role of FER and highlighting its potential as a novel therapeutic target for metastatic ovarian cancer.
Keywords: FER; GAB1; MET; ovarian cancer; tyrosine phosphorylation.
© 2016 Fan et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
Similar articles
-
FER mediated HGF-independent regulation of HGFR/MET activates RAC1-PAK1 pathway to potentiate metastasis in ovarian cancer.Small GTPases. 2020 May;11(3):155-159. doi: 10.1080/21541248.2017.1379931. Epub 2018 Jan 24. Small GTPases. 2020. PMID: 29099290 Free PMC article.
-
Spatial regulation of signaling by the coordinated action of the protein tyrosine kinases MET and FER.Cell Signal. 2018 Oct;50:100-110. doi: 10.1016/j.cellsig.2018.06.006. Epub 2018 Jun 18. Cell Signal. 2018. PMID: 29920310 Free PMC article.
-
The multisubstrate adapter Gab1 regulates hepatocyte growth factor (scatter factor)-c-Met signaling for cell survival and DNA repair.Mol Cell Biol. 2001 Aug;21(15):4968-84. doi: 10.1128/MCB.21.15.4968-4984.2001. Mol Cell Biol. 2001. PMID: 11438654 Free PMC article.
-
Structure, biosynthesis and biochemical properties of the HGF receptor in normal and malignant cells.EXS. 1993;65:131-65. EXS. 1993. PMID: 8380735 Review.
-
HGF/MET signaling in ovarian cancer.Curr Mol Med. 2008 Sep;8(6):469-80. doi: 10.2174/156652408785747933. Curr Mol Med. 2008. PMID: 18781954 Review.
Cited by
-
HGF-mediated crosstalk between cancer-associated fibroblasts and MET-unamplified gastric cancer cells activates coordinated tumorigenesis and metastasis.Cell Death Dis. 2018 Aug 29;9(9):867. doi: 10.1038/s41419-018-0922-1. Cell Death Dis. 2018. PMID: 30158543 Free PMC article.
-
LncRNA DLEU1 facilitates the progression of oral squamous cell carcinoma by miR-126-5p/GAB1 axis.Kaohsiung J Med Sci. 2022 Oct;38(10):950-959. doi: 10.1002/kjm2.12572. Epub 2022 Aug 30. Kaohsiung J Med Sci. 2022. PMID: 36039933 Free PMC article.
-
Targeting FER Kinase Inhibits Melanoma Growth and Metastasis.Cancers (Basel). 2019 Mar 24;11(3):419. doi: 10.3390/cancers11030419. Cancers (Basel). 2019. PMID: 30909648 Free PMC article.
-
Rac1, A Potential Target for Tumor Therapy.Front Oncol. 2021 May 17;11:674426. doi: 10.3389/fonc.2021.674426. eCollection 2021. Front Oncol. 2021. PMID: 34079763 Free PMC article. Review.
-
Protein tyrosine phosphatase receptor type R (PTPRR) antagonizes the Wnt signaling pathway in ovarian cancer by dephosphorylating and inactivating β-catenin.J Biol Chem. 2019 Nov 29;294(48):18306-18323. doi: 10.1074/jbc.RA119.010348. Epub 2019 Oct 25. J Biol Chem. 2019. PMID: 31653698 Free PMC article.
References
-
- Ahn J, Truesdell P, Meens J, Kadish C, Yang X, Boag AH, Craig AW. 2013. Fer protein-tyrosine kinase promotes lung adenocarcinoma cell invasion and tumor metastasis. Mol Cancer Res 11: 952–963. - PubMed
-
- Albeck JG, Brugge JS. 2011. Uncovering a tumor suppressor for triple-negative breast cancers. Cell 144: 638–640. - PubMed
-
- Arend RC, Londono-Joshi AI, Straughn JM Jr, Buchsbaum DJ. 2013. The Wnt/β-catenin pathway in ovarian cancer: a review. Gynecol Oncol 131: 772–779. - PubMed
-
- Ayhan A, Ertunc D, Tok EC, Ayhan A. 2005. Expression of the c-Met in advanced epithelial ovarian cancer and its prognostic significance. Int J Gynecol Cancer 15: 618–623. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous