

Long non-coding RNAs are major contributors to transcriptome changes in sunflower meiocytes with different recombination rates
- PMID: 27401977
- PMCID: PMC4940957
- DOI: 10.1186/s12864-016-2776-1
Long non-coding RNAs are major contributors to transcriptome changes in sunflower meiocytes with different recombination rates
Abstract
Background: Meiosis is a form of specialized cell division that marks the transition from diploid meiocyte to haploid gamete, and provides an opportunity for genetic reassortment through recombination. Experimental data indicates that, relative to their wild ancestors, cultivated sunflower varieties show a higher recombination rate during meiosis. To better understand the molecular basis for this difference, we compared gene expression in male sunflower meiocytes in prophase I isolated from a domesticated line, a wild relative, and a F1 hybrid of the two.
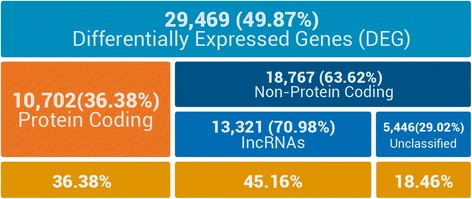
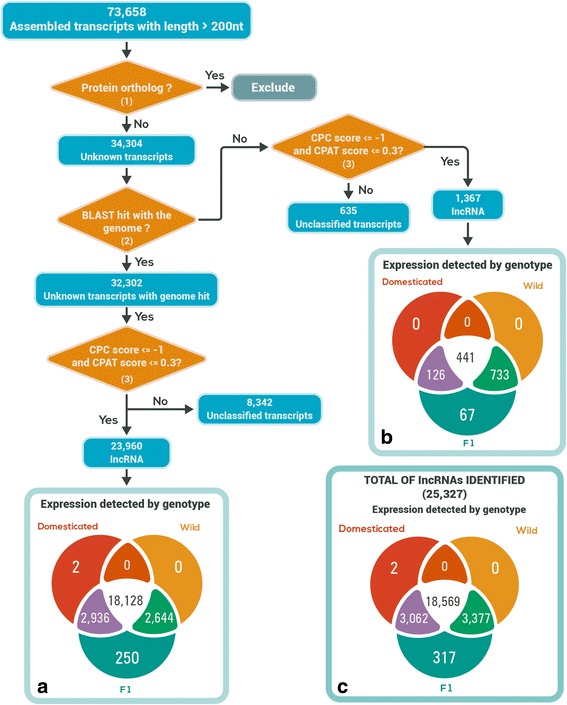
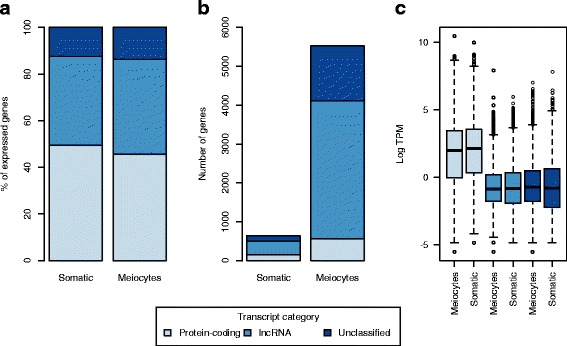
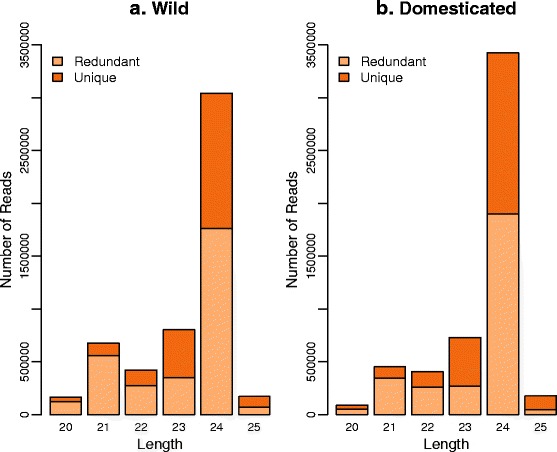
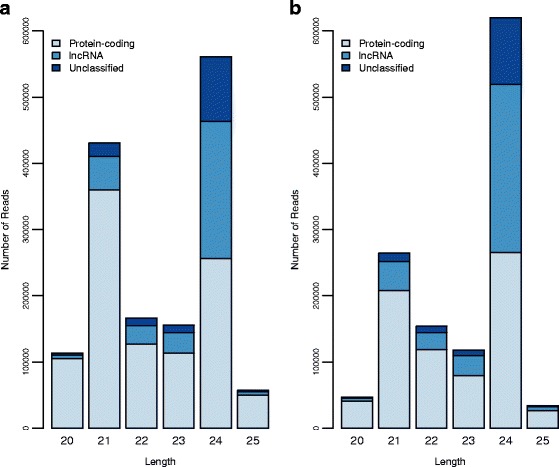
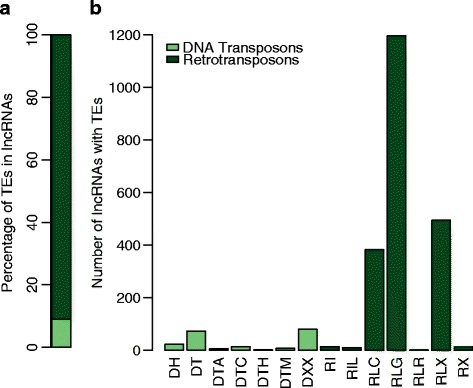
Results: Of the genes that showed differential expression between the wild and domesticated genotypes, 63.62 % could not be identified as protein-coding genes, and of these genes, 70.98 % passed stringent filters to be classified as long non-coding RNAs (lncRNAs). Compared to the sunflower somatic transcriptome, meiocytes express a higher proportion of lncRNAs, and the majority of genes with exclusive expression in meiocytes were lncRNAs. Around 40 % of the lncRNAs showed sequence similarity with small RNAs (sRNA), while 1.53 % were predicted to be sunflower natural antisense transcripts (NATs), and 9.18 % contained transposable elements (TE). We identified 6895 lncRNAs that are exclusively expressed in meiocytes, these lncRNAs appear to have higher conservation, a greater degree of differential expression, a higher proportion of sRNA similarity, and higher TE content relative to lncRNAs that are also expressed in the somatic transcriptome.
Conclusions: lncRNAs play important roles in plant meiosis and may participate in chromatin modification processes, although other regulatory functions cannot be excluded. lncRNAs could also be related to the different recombination rates seen for domesticated and wild sunflowers.
Keywords: Homologous recombination; Meiosis; Prophase I; RNA-seq; Transcriptomics; lncRNA; miRNA.
Figures
References
-
- Armstrong SJ, Franklin FCH, Jones GH. A meiotic time-course for Arabidopsis thaliana. Sex Plant Reprod. 2003;16:141–9. doi: 10.1007/s00497-003-0186-4. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
